Jane Flint - Principles of Virology
Здесь есть возможность читать онлайн «Jane Flint - Principles of Virology» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.
- Название:Principles of Virology
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Principles of Virology: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Principles of Virology»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Volume I: Molecular Biology
Volume II: Pathogenesis and Control
Principles of Virology, Fifth Edition
Principles of Virology — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Principles of Virology», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
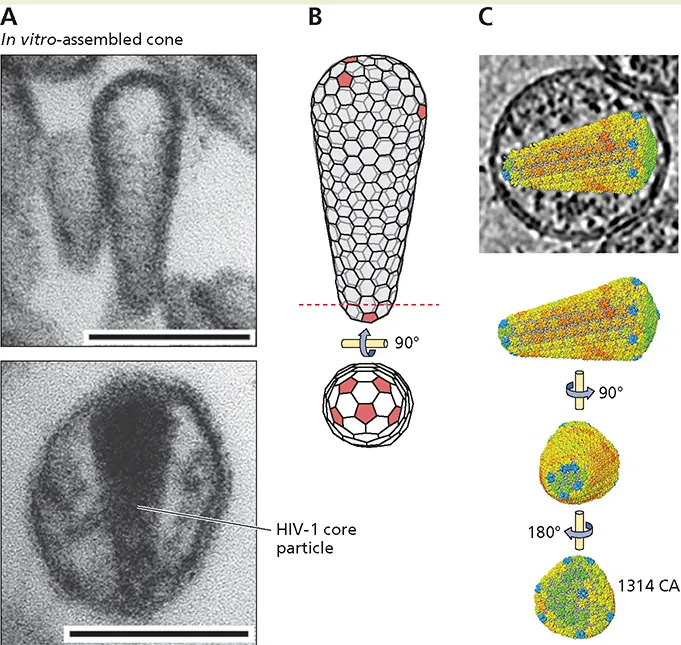
Li S, Hill CP, Sundquist WI, Finch JT. 2000. Image reconstructions of helical assemblies of the HIV-1 CA protein. Nature 407:409–413.
Zhao G, Perilla JR, Yufenyuy EL, Meng X, Chen B, Ning J, Ahn J, Gronenborn AM, Schulten K, Aiken C, Zhang P. 2013. Mature HIV-1 capsid structure by cryo-electron microscopy and all-atom molecular dynamics. Nature 497:643–646.
Mattei S, Glass B, Hagen WJH, Kräusslich H-G, Briggs JAG. 2016. The structure and flexibility of conical HIV-1 capsids determined within intact virions. Science 354:1434–1437.
Other Capsid Architectures
As noted previously, capsids with icosahedral or helical symmetry are characteristic of the majority of virus particles described to date. Nevertheless, these architectures are not universal, and the capsids of even some relatively small viruses can be constructed according to an alternative design. This property is exemplified by the capsids of some retroviruses.
The capsid of all retroviruses surrounds a nucleoprotein that contains the diploid (+) strand RNA genome. The capsid is surrounded by the viral matrix (M) protein, which is encased in the viral envelope. However, retroviral capsids may be spherical, cylindrical, or conical, the shape exhibited by capsids of human immunodeficiency virus type 1 and other lentiviruses ( Fig. 4.18Aand B). These capsids are built from a single capsid (CA) protein, which can form both pentamers and hexamers. The odd appearance of the human immunodeficiency virus type 1 capsid might suggest that it represents an exception to the geometric rules that dictate the viral architectures described in previous sections. However, this is not the case: this capsid can be described by a fullerene cone model that combines principles of both icosahedral and helical symmetry. In this model ( Box 4.6), which has been confirmed by cryo-electron tomography of intact virus particles, a closed structure is formed using 12 pentamers, just as in an icosahedral capsid. However, pentamers are not spaced at regular intervals throughout the structure. Rather, they are restricted to the terminal caps and separated by spirals (a variant of helical symmetry) of CA hexamers that form the body of the cone.
Packaging the Nucleic Acid Genome
A definitive property of a virion is the presence of a nucleic acid genome. Incorporation of the genome requires its discrimination from a large population of cellular nucleic acid. This packaging process is described in Chapter 13. The volumes of closed capsids are finite. Consequently, accommodation of viral genomes necessitates a high degree of condensation and compaction. A simple analogy illustrates vividly the scale of this problem; packing of the ~150-kbp DNA genome of herpes simplex virus type 1 into the viral capsid is equivalent to stuffing some 10 ft of 22 American gauge wire (diameter, 0.644 mm) into a tennis ball. Such confinement of the genome can result in high internal pressure, equivalent to that generated in locomotive steam engines, some 18 and 25 atm within herpes simplex virus type 1 and phage capsids, respectively. Such pressure provides the force that powers ejection of DNA genomes. Packaging of nucleic acids is an intrinsically unfavorable process because of the highly constrained conformation imposed on the genome. In some cases, the force required to achieve packaging is provided, at least in part, by specialized viral proteins that harness the energy released by hydrolysis of ATP to drive the insertion of DNA. In many others, the binding of viral RNA or DNA genomes to capsid proteins appears to provide sufficient energy. The latter interactions also help to neutralize the negative charge of the sugar-phosphate backbone, a prerequisite for close juxtaposition of genome sequences.
We possess relatively little information about the organization of genomes within viral particles: nucleic acids or protein-nucleic acid assemblies are not visible in the majority of high-resolution structural studies reported. This limitation indicates that the genomes or internal structures lack the symmetry of the capsid, do not adopt the same conformation in every viral particle, or both. Nevertheless, three mechanisms for condensing and organizing nucleic acid molecules within capsids can be distinguished and are described in the following sections.
Direct Contact of the Genome with a Protein Shell
In the simplest arrangement, the nucleic acid makes direct contact with the protein(s) that forms the protective shell of the virus particle. Proteins on the inner surfaces of the icosahedral capsids of many small RNA viruses interact with the viral genome. As we have seen, the interior surface of the poliovirus capsid can be described in detail. Nevertheless, we possess no structural information about the arrangement of the RNA genome, for the nucleic acid is not visible in the X-ray structure. However, the genome of the porcine picornavirus Seneca Valley virus has been visualized by this method (albeit at low resolution) ( Fig. 4.19A). Much of the RNA genome forms an outer layer in which it makes extensive contact with the inner surface of the capsid. Highly ordered RNA genomes are also present in T = 3 nodaviruses, such as Flock house virus, in which an outer decahedral cage of ordered RNA surrounds additional rings (Fig 4.19B). More recently, higher-resolution views (sufficient to observe some bases in the RNA!) of a small viral RNA genome have been obtained using cryo-EM reconstruction without imposition of any symmetry ( Box 4.7). This approach is likely to be more widely applicable.
Use of the same protein or proteins both to condense the genome and to build a capsid allows efficient utilization of limited genetic capacity. It is therefore an advantageous arrangement for viruses with small genomes. However, this mode of genome packing is also characteristic of some larger viruses, notably rotaviruses and herpesviruses. The genome of rotaviruses comprises 11 segments of double-stranded RNA located within the innermost of the three protein shells of the particle. Remarkably, as much as 80% of the RNA genome appears highly ordered within the core, with strong elements of icosahedral symmetry ( Fig. 4.19B).
One of the most surprising properties of the large herpesviral capsid is the absence of internal proteins associated with viral DNA: despite intense efforts, no such core proteins have been identified, and the viral genome has not yet been visualized. In contrast, cryo-EM has allowed visualization of the large, double-stranded DNA genome of bacteriophage T4, which is organized in closely apposed, concentric layers ( Fig. 4.20A). This arrangement illustrates graphically the remarkably dense packing needed to accommodate such large viral DNA genomes in closed structures of fixed dimensions. This type of organization must require neutralization of the negative charges of the sugar-phosphate backbone. Neutralization might be accomplished by proteins that form the inner surface of the capsid, or by the incorporation of small, basic peptides made by the host cell, such as spermine and spermidine.
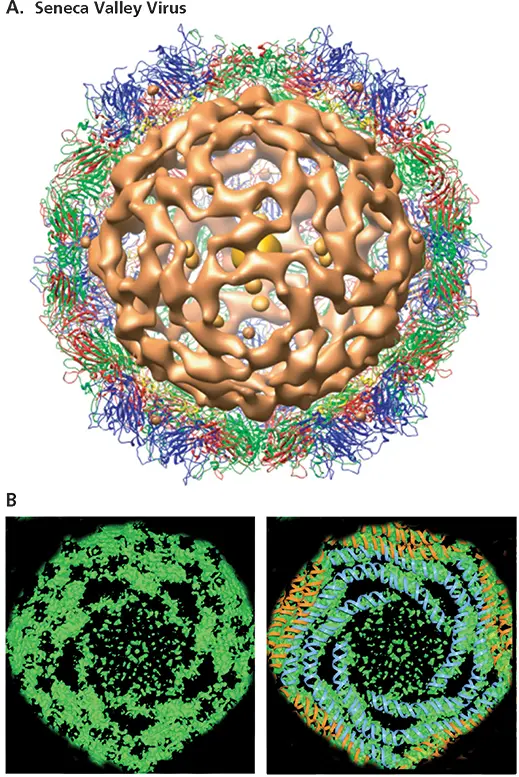
Figure 4.19 Ordered RNA genomes in small and large icosahedral virus particles. (A)The 20-Å X-ray crystal structure of the picornavirus Seneca Valley virus viewed down a twofold axis of icosahedral symmetry, showing the density ascribed to the RNA genome (brown). The structural proteins are colored as in Fig. 4.12and 4.13: VP1 (blue), VP2 (green), VP3 (red), and VP4 (yellow). (B)Outer layer of the double-stranded, segmented RNA genome of the rotavirus bluetongue virus observed at 6.5-Å resolution by X-ray crystallography of viral cores. The electron density of this layer of RNA (green) from maps averaged between two closely related serotypes is shown with A-form duplex RNA modeled into the rods of density. These RNA spirals represent some 80% of the >19-kbp genome. Reprinted from Gouet P et al. 1999. Cell 97:-481–490, with permission. Courtesy of D.I. Stuart, University of Oxford.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Principles of Virology»
Представляем Вашему вниманию похожие книги на «Principles of Virology» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Principles of Virology» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.