Jane Flint - Principles of Virology
Здесь есть возможность читать онлайн «Jane Flint - Principles of Virology» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.
- Название:Principles of Virology
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Principles of Virology: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Principles of Virology»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Volume I: Molecular Biology
Volume II: Pathogenesis and Control
Principles of Virology, Fifth Edition
Principles of Virology — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Principles of Virology», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Some viruses multiply in cells without causing obvious cytopathic effects. For example, many members of the families Arenaviridae , Paramyxoviridae , and Retroviridae do not cause obvious damage to cultured cells. Infection by such viruses must therefore be assessed using alternative methods, as described in “Assay of Viruses” below.
Embryonated Eggs
Before the advent of cell culture, many viruses were propagated in embryonated chicken eggs ( Fig. 2.6). At 5 to 14 days after fertilization, a hole is drilled in the shell and virus is injected into the site appropriate for its replication. This method of virus propagation is now routine only for influenza virus. The robust yield of this virus from chicken eggs has led to their widespread use in research laboratories and for vaccine production.
Laboratory Animals
In the early 1900s, when viruses were first isolated, freezers and cell cultures were not available, and it was necessary to maintain virus stocks by continuous passage from animal to animal. This practice not only was inconvenient but also, as we shall see, led to the selection of viral mutants (Volume II, Chapter 7). For example, monkey-to-monkey intracerebral passage of poliovirus selected a mutant that could no longer infect chimpanzees by the oral route, the natural means of infection.
Although cell culture has supplanted animals for propagating most viruses, experimental infection of laboratory animals has always been, and will continue to be, obligatory for studying the processes by which viruses cause disease. The study in monkeys of poliomyelitis, the paralytic disease caused by poliovirus, led to an understanding of the basis of this disease and was instrumental in the development of a successful vaccine. Similarly, the development of vaccines against hepatitis B virus would not have been possible without experimental studies with chimpanzees. Understanding how the immune system or any complex organ reacts to a virus cannot be achieved without research on living animals. The development of viral vaccines, antiviral drugs, and diagnostic tests for veterinary medicine has also benefited from research on diseases in laboratory animals. Despite their utility, it must be appreciated that all animal models are surrogates for the events that occur during viral infections of humans.
Assay of Viruses
There are two main types of assay for detecting viruses: biological and physical. Because viruses were first recognized by their infectivity, the earliest assays focused on this most sensitive and informative property. However, biological assays such as the plaque assay and end-point titration methods do not detect noninfectious particles. In contrast, all particles are accounted for with physical assays such as electron microscopy or by immunological methods. Knowledge of the number of noninfectious particles is useful for assessing the quality of a virus preparation.
Measurement of Infectious Units
One of the most important procedures in virology is measuring the virus titer, the concentration of infectious virus particles in a sample. This parameter is determined by inoculating serial dilutions of virus into host cell cultures, chicken embryos, or laboratory animals and monitoring for evidence of virus multiplication. The response may be quantitative (as in assays for plaques, fluorescent foci, infectious centers, or abnormal growth and morphology) or all-or-none, in which the presence or absence of infection is measured (as in an end-point dilution assay). Please note that “titer” is not a verb.
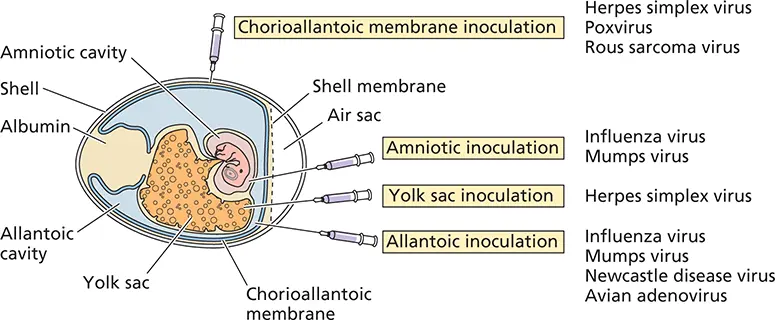
Figure 2.6 Growth of viruses in embryonated eggs. The cutaway view of an embryonated chicken egg shows the different routes by which viruses are inoculated into eggs and the distinct compartments in which particular viruses may propagate.
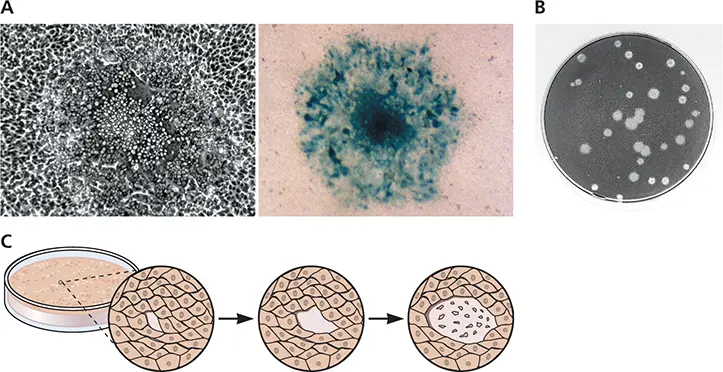
Figure 2.7 Plaques formed by different animal viruses. (A)Photomicrograph of a single plaque formed by pseudorabies virus in bovine kidney cells. Shown are unstained cells (left) and cells stained with the chromogenic substrate X-Gal (5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside), which is converted to a blue compound by the product of the lacZ gene carried by the virus (right). Courtesy of B. Banfield, Princeton University. (B)Plaques formed by poliovirus on human HeLa cells stained with crystal violet. (C)Illustration of the sequential spread of a cytopathic virus from an initial infected cell to neighboring cells, resulting in a plaque.
Plaque Assay
The measurement of virus titers by plaque assay was first developed for bacteriophages by d’Herelle in 1917 and then modified for animal viruses by Renato Dulbecco in 1952. In this procedure, monolayers of cultured cells are incubated with a preparation of virus to allow adsorption to cells. After removal of the inoculum, the cells are covered with nutrient medium containing a supplement, most commonly agar, which forms a gel. When the original infected cells release new progeny particles, the gel restricts their spread to neighboring uninfected cells. As a result, each infectious particle produces a circular zone of infected cells, a plaque. If the infected cells are damaged, the plaque can be distinguished from the surrounding monolayer. In time, the plaque becomes large enough to be seen with the naked eye ( Fig. 2.7). Only viruses that cause visible damage of cultured cells can be assayed in this way. A movie that depicts the microscopic development of a plaque can be found at this link: http://bit.ly/Virology_VZVGFP.
For the majority of animal viruses, there is a linear relationship between the number of infectious particles and the plaque count ( Fig. 2.8). One infectious particle is therefore sufficient to initiate infection, and the virus is said to infect cells with one-hit kinetics. Some examples of two-hit kinetics, in which two different types of virus particle must infect a cell to ensure replication, have been recognized. An example is the genomes of some (+) strand RNA viruses of plants that consist of two RNA molecules which are encapsidated separately. Both RNAs are required for infectivity. The dose-response curve in plaque assays for these viruses is therefore parabolic rather than linear ( Fig. 2.8).
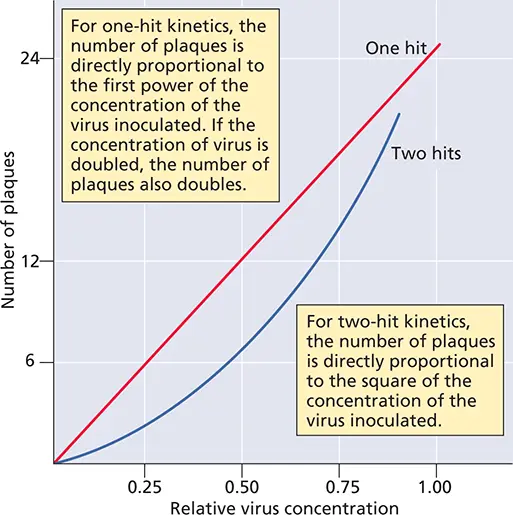
Figure 2.8 The dose-response curve of the plaque assay. The number of plaques produced by a virus with one-hit kinetics (red) or two-hit kinetics (blue) is plotted against the relative concentration of the virus. In two-hit kinetics, there are two classes of uninfected cells, those receiving one particle and those receiving none. The Poisson distribution can be used to determine the proportion of cells in each class: they are e–m and me–m ( Box 2.12). Because one particle is not sufficient for infection, P (0) = e–m (1 + m ). At a very low multiplicity of infection, this equation becomes P (i) = (1/2) m 2(where i = infection), which gives a parabolic curve.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Principles of Virology»
Представляем Вашему вниманию похожие книги на «Principles of Virology» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Principles of Virology» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.