George Acquaah - Principles of Plant Genetics and Breeding
Здесь есть возможность читать онлайн «George Acquaah - Principles of Plant Genetics and Breeding» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Principles of Plant Genetics and Breeding
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Principles of Plant Genetics and Breeding: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Principles of Plant Genetics and Breeding»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Organizes topics to reflect the stages of an actual breeding project Incorporates the most recent technologies in the field, such as CRSPR genome edition and grafting on GM stock Includes numerous illustrations and end-of-chapter self-assessment questions, key references, suggested readings, and links to relevant websites Features a companion website containing additional artwork and instructor resources
offers researchers and professionals an invaluable resource and remains the ideal textbook for advanced undergraduates and graduates in plant science, particularly those studying plant breeding, biotechnology, and genetics.
Principles of Plant Genetics and Breeding — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Principles of Plant Genetics and Breeding», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
The 28→38 apomictic transfer pathway
This pathway has been described only once and has been little examined or discussed in the literature. Consequently, some detail regarding the results in this research will be presented. In 1958, Dr. M. Borovsky, Institute of Agriculture, Kishinev, Moldova, performed a series of hybridizations between a diploid popcorn line identified as Risovaia 645 and a sexual diploid (2n = 2x = 36) T. dactyloides clone with the first maize‐ Tripsacum hybrids being generated in 1960 (Borovsky 1966; Borovsky and Kovarsky 1967). The F 1hybrids generated from the experiments possessed 28 chromosomes (10Mz + 18Tr). The F 1plants were completely male sterile and were highly seed sterile. Backcrossing with diploid maize identified that some of the F 1hybrids were approximately 1–1.5% seed fertile and resulted in the production of progeny possessing 28 chromosomes (10Mz + 18Tr) and 38 chromosomes (20Mz + 18Tr). When the F 1was backcrossed to the Tripsacum parent, the fertile F 1's generated progeny with 28 chromosomes (10Mz + 18Tr) and 46 chromosomes (10Mz − 18Tr + 18Tr). The complete set of backcrosses with maize and Tripsacum resulted in a ratio of approximately 10 (28‐chromosome plants) to one (38‐ or 46‐chromosome plant). Phenotypic observations suggested that the 28‐chromosome progeny were not different from their 28‐chromosome parent while the 38‐ and 46‐chromosome progeny were clearly different. In addition, some seed generated by the 28‐chromosome F 1's were polyembryonic. Additional evaluations on the 28‐chromosome F 1and its 28‐chromosome progeny suggested that these F 1plants and their progeny were apomictic. This early, non‐replicated experiment is to date the only report where a 28‐chromosome F 1hybrid was maintained by apomixis. Polyembryony was noted, and a diploid sexual Tripsacum was used to generate the interspecific hybrid.
The 46→56→38 non‐apomictic pathway
This sexual or non‐apomictic pathway, as discussed by Harlan and deWet, is believed to offer the greatest opportunity for Tripsacum introgression into maize and represents results of an early attempt to transfer apomixis to maize (Petrov et al. 1979, 1984). In this pathway, a diploid maize (2n = 2x = 20Mz) is crossed with a tetraploid Tripsacum resource (2n = 4x = 72Tr). The resultant F 1hybrid possesses 10Mz + 36Tr chromosomes. To date, published reports indicate all of these hybrids are pollen sterile and vary considerably in their levels of seed fertility. In this particular pathway, when the 46‐chromosome F 1is backcrossed to diploid maize, 56‐chromosome individuals result following fertilization of an unreduced egg. As in the prior pathways, this result is generated by a 2n + n mating event. Offspring of the 56‐chromosome individual following a second backcross to maize generally possess 38 chromosomes (20Mz + 18Tr) and resemble those discussed in the 28→38→20 pathway above. The generation of progeny with 38 chromosomes is the result of meiosis in the developing megaspore. In this instance, the maize and Tripsacum complements pair with their homologous sets (Mz–Mz, Tr–Tr). Following a complete occurrence of meiosis I and II divisions, the result is a reduced egg having 10Mz + 18Tr chromosomes, which when backcrossed by a diploid maize, results in progeny having 20Mz + 18Tr chromosomes. Almost exclusively, the 38‐chromosome individuals no longer express any level or form of an apomictic reproductive mechanism, and subsequent backcrossing to maize results in the recovery of individuals possessing 20Mz and a varying number of Tripsacum chromosomes. Upon backcrossing, the 38‐chromosome individuals behave in an identical manner to their 28‐chromosome cousins represented in the 28→38→20 pathway. Generally, Tripsacum introgression by homoeologous pairing and recombination does not occur and genetic transfer of Tripsacum genes to maize is not accomplished.
The 46→56→38 apomictic transfer pathway
Though not addressed in Harlan and de Wet's 1977 research, this pathway is similar enough and worthwhile to discuss with regard to its relevance to apomixis. First published by Petrov and colleagues as early as 1979, and replicated in similar style by others, a diploid or tetraploid maize line is pollinated by a tetraploid, apomictic T. dactyloides clone (Petrov et al. 1979, 1984). If a diploid maize line is utilized, the resultant F 146‐chromosome hybrid possesses 10Mz and 36Tr chromosomes. Upon backcrossing with diploid maize, both apomictic 46‐chromosome and 56‐chromosome (20Mz + 36Tr) individuals can be obtained. The 46‐chromosome offspring are products of apomixis. The 56‐chromosome offspring are products of an unreduced egg being fertilized by the diploid maize pollen source, another 2n + n mating event. Often, these individuals exhibit polyembryony that results in the generation of “twins” being obtained from a single seed ( Figure B6.3). In some instances, these polyembryonic events give rise to 46–46 pairs of twins (each apomictic clones of the other), 46–56 “twins,” one arising from a unfertilized reduced egg, the other arising from a fertilized unreduced egg; and in some instances varying combinations of 46‐46‐46 or 46‐46‐56 triplets. Typically, as seedlings, the 56‐chromosome individuals are more vigorous than their 46‐chromosome sibs.
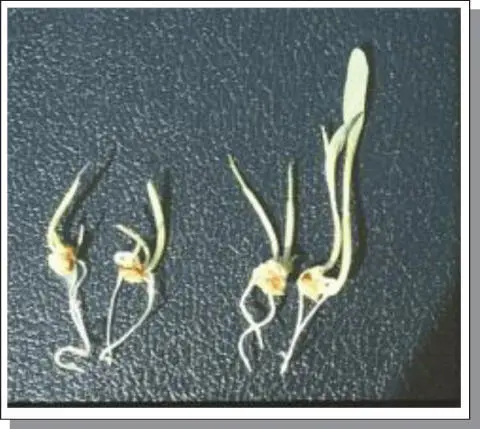
Figure B6.3Polyembryony expression in germinating seed of an apomictic 46‐chromosome F 1maize‐ Tripsacum hybrid. Note in the pairs, one seedling is often larger and more vigorous than its sib. The larger sib of the pair is often the product of a 2n + n mating event.
Backcrossing the 46‐chromosome individuals by maize, repeats the above cycle. Upon backcrossing the 56‐chromsome individuals with maize, three types of progeny can be observed. Typically, progeny having 56 chromosomes are generated. However, in some instances, 2n + n matings occur, giving rise to individuals possessing 66 chromosomes (30Mz + 36Tr). Occasionally, a reduced egg will be generated and may or may not be fertilized by the available maize pollen. In rare instances of non‐fertilization, a 28‐chromosome individual is generated (10Mz + 18Tr). In instances whereby the maize pollen fertilizes the reduced egg, 38‐chromosome individuals are obtained (20Mz + 18Tr). Generally, individuals possessing 38 chromosomes, rather than 28 chromosomes, are the most common product. What is unique about this pathway is that occasionally, the 38‐chromosome individuals retain all the elements of apomixis which were present in the Tripsacum paternal parent and the F 1and BC1 individuals. The retention of apomixis to this 38‐chromosome level has been well documented and repeated in several laboratories (Petrov et al. 1979, 1984; Leblanc et al. 1996; Kindiger and Sokolov 1997). In addition, the occurrence of 2n + n matings, polyembryony and variation in apomixis expression is quite similar to that found in apomictic Tripsacum (Kindiger et al. 1996a).
Following the generation and confirmation of apomictic 38‐chromsome individuals (20Mz + 18Tr), it is apparently a difficult and uncommon occurrence to generate and maintain apomixis in backcross generations that have fewer Tripsacum chromosomes. Only one report has been published where apomictic individuals possessing only 9Tr chromosomes were obtained (Kindiger et al. 1996b). Generally, by 2n + n mating events, the 38‐chromosome individuals produce only apomictic 38‐chromosome progeny and 48‐chromsome progeny. Backcrossing the 48‐chromosome individuals results in 48‐chromosome apomictics and 58‐chromosome apomictics. This accumulation of maize genomes continues until a point is achieved where the additional maize genomes eventually shift the individual from an apomictic reproductive mechanism to a traditional sexual mode of reproduction, whence, apomixis is never again attained. This commonly occurs when five or six doses (50–60 maize chromosomes) are present. The result of a 78‐chromosome individual (60Mz + 18Tr) losing apomixis is the return of meiosis and a highly seed‐sterile individual producing an array of highly maize‐like aneuploids with a random set of Tripsacum chromosomes. Backcrossing these individuals, that are typically pollen sterile, generally results in the recovery of diploid maize lines with or without any Tr chromosomes. To date, the apomictic maize‐ Tripsacum line possessing 39 chromosomes (30Mz + 9Tr) represents the most advanced level of apomixis transfer to maize. An array of various ear types generated from a series of maize‐ Tripsacum hybrids is provided in Figure B6.4.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Principles of Plant Genetics and Breeding»
Представляем Вашему вниманию похожие книги на «Principles of Plant Genetics and Breeding» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Principles of Plant Genetics and Breeding» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.