Michael Begon - Ecology
Здесь есть возможность читать онлайн «Michael Begon - Ecology» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Ecology
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Ecology: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Ecology»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
– now in full colour – offers students and practitioners a review of the ecological sciences.
The previous editions of this book earned the authors the prestigious ‘Exceptional Life-time Achievement Award’ of the British Ecological Society – the aim for the fifth edition is not only to maintain standards but indeed to enhance its coverage of Ecology.
In the first edition, 34 years ago, it seemed acceptable for ecologists to hold a comfortable, objective, not to say aloof position, from which the ecological communities around us were simply material for which we sought a scientific understanding. Now, we must accept the immediacy of the many environmental problems that threaten us and the responsibility of ecologists to play their full part in addressing these problems. This fifth edition addresses this challenge, with several chapters devoted entirely to applied topics, and examples of how ecological principles have been applied to problems facing us highlighted throughout the remaining nineteen chapters.
Nonetheless, the authors remain wedded to the belief that environmental action can only ever be as sound as the ecological principles on which it is based. Hence, while trying harder than ever to help improve preparedness for addressing the environmental problems of the years ahead, the book remains, in its essence, an exposition of the
of ecology. This new edition incorporates the results from more than a thousand recent studies into a fully up-to-date text.
Written for students of ecology, researchers and practitioners, the fifth edition of
is anessential reference to all aspects of ecology and addresses environmental problems of the future.
Ecology — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Ecology», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
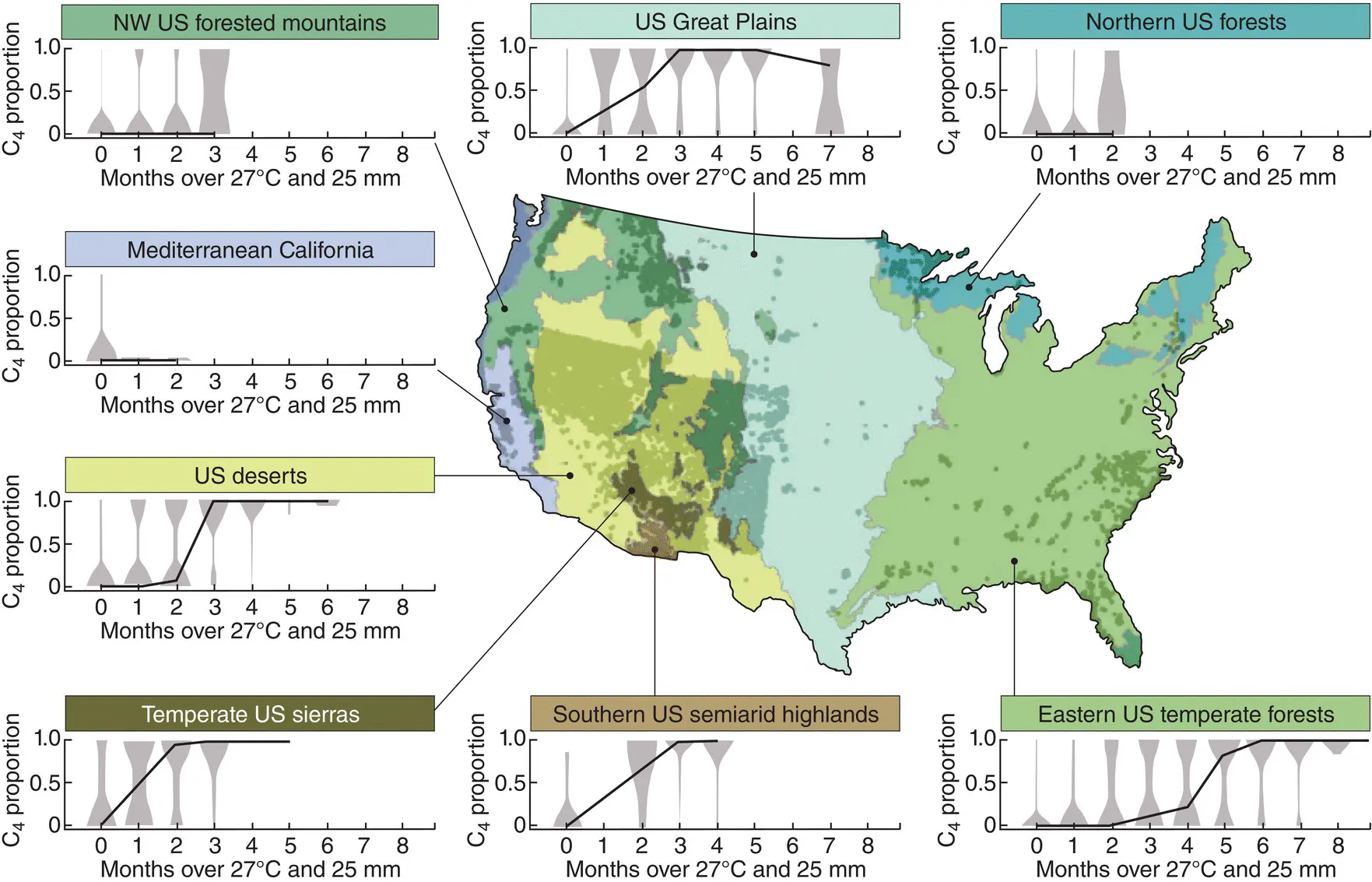
Figure 3.17 Effects of temperature and precipitation on the proportional contributions of C 3 and C 4 grasses to the floras of various regions of the USA, as indicated.Data were collected from sampling plots within each region, with their location marked as dots on the map, and these data are shown as symmetrical ‘density curves’, associated with the number of months at each location where conditions exceeded the estimated temperature‐precipitation threshold (to the nearest month). The solid line is the predicted median proportion derived from a ‘quantile regression’ based on the temperature–precipitation threshold.
Source : After Griffith et al . (2015).
the CAM pathway
Plants with a CAM pathway also use PEP carboxylase with its strong power of concentrating CO 2. (The system is now known in a wide variety of families, not just the Crassulaceae.) In contrast to C 3and C 4plants, though, CAM plants open their stomata and fix CO 2at night (as malic acid). During the daytime the stomata are closed and the CO 2is released within the leaf and fixed by RuBisCO. However, because the CO 2is then at a high concentration within the leaf, photorespiration is prevented, just as it is in plants using the C 4pathway. Plants using the CAM photosynthetic pathway have obvious advantages when water is in short supply, because their stomata are closed during the daytime when evaporative forces are strongest. This appears to be a highly effective means of water conservation – water use efficiency for CAM plants is estimated to be around three times greater than for C 4plants and more than six times greater than for C 3plants (Borland et al ., 2009) – but CAM species have not come to inherit the earth. One cost to CAM plants is the problem of storing the malic acid that is formed at night: most CAM plants are succulents with extensive water‐storage tissues that cope with this problem. In general, CAM plants are found in arid environments where strict stomatal control of daytime water is vital for survival (desert succulents), and in habitats where CO 2is in short supply during the daytime, for example in some submerged aquatic plants, and in photosynthetic organs that lack stomata (e.g. the aerial photosynthetic roots of orchids).
APPLICATION 3.2 Turning to CAM crops
The high water use efficiency of CAM plants makes them excellent candidates for cultivation over areas where rainfall is too little or evapotranspiration too high for the cultivation of C 3or C 4crops. Such areas may well expand in future. Examples include the production of pineapple, Ananas comosus , for consumption, of Opuntia spp. for animal fodder, of sisal, Agave sisalana , for fibre, and of Agave tequilana for the alcoholic beverages tequila and mescal (Borland et al ., 2009). In addition, species of Agave are grown for both liquid and gaseous biofuels and indeed other products such as enzymes and lactic and succinic acid (Perez‐Pimienta et al ., 2017). Current quantities grown are small compared with the most common alternatives based around corn, sugar cane and wheat, and most of this is in Mexico. However, the much greater productivity of Agave ( Figure 3.18), and the high water use efficiency it derives from its CAM metabolism, highlights its great potential for the future.
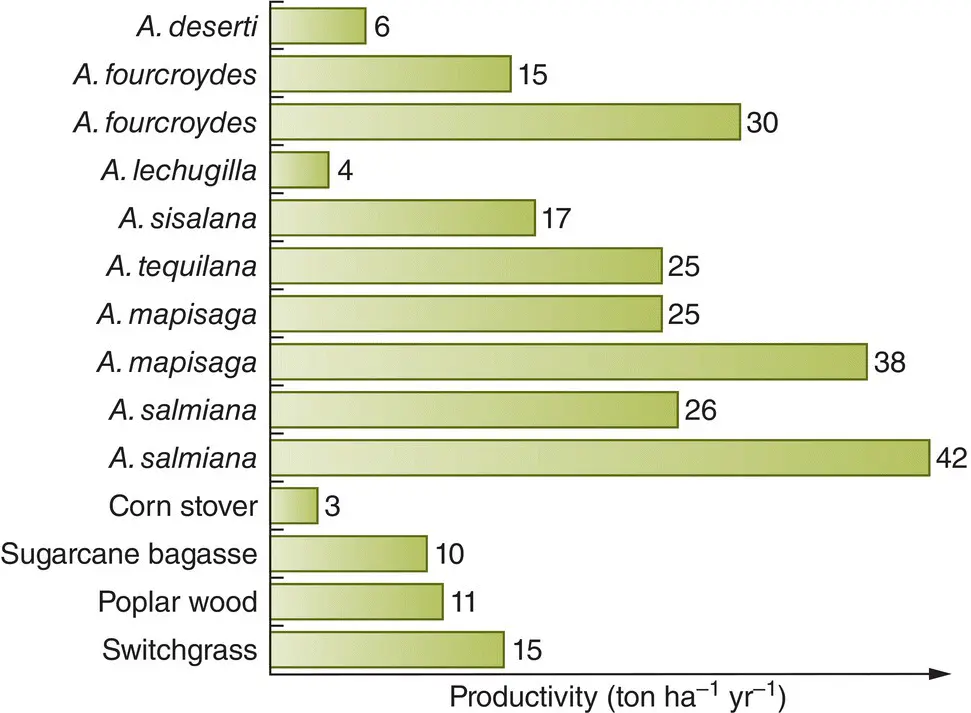
Figure 3.18 Agave species (CAM plants) exhibit high productivity due to their high water use efficiency.Comparison of the productivities of various species of Agave (double entries reflect separate studies) and four selected bioenergy feedstocks (raw products for the production of biofuels). Corn stover consists of the remains of plants left in the field after corn ( Zea mays ) has been harvested. Sugarcane bagasse is the fibrous matter than remains after sugarcane ( Saccharum spp.) has been crushed to extract its juice.
Source : After Perez‐Pimienta et al . (2017).
APPLICATION 3.3 Genetic engineering of CCMs into crops
Another way to exploit the potential of CAM would be to engineer its molecular machinery into C 3plants (Borland et al ., 2014). Indeed there is great interest in general in engineering a wider range of CCMs, taken from C 4plants and from microbes, into commercial crops. Complex metabolic alternatives, such as CAM, typically require a whole suite of co‐adapted changes, and implementing them will therefore be a massive challenge. However, promising results may be obtained from less ambitious approaches. Figure 3.19, for example, illustrates the results when the photosynthetic rate of a standard variety of soybean, Glycine max , was compared with that of a transgenic variety that had been transformed to express the ictB gene taken from cyanobacteria, where it plays a crucial role in carbon uptake and is essential for their survival at anything other than very high CO 2concentrations. In both varieties, photosynthetic rates increased with the concentrations of CO 2in plant tissues but levelled off at higher concentrations. However, these rates, and especially the maximum rate, were significantly higher in the transgenic variety ( Figure 3.19). Crucially, this difference translated into significant improvements in yield in the transgenic variety. As the authors of the study conclude, work such as this indicates that even single genes can contribute to the enhancement of yield in a major commodity crop ‘… and point to the significant role that biotechnological approaches to increasing photosynthetic efficiency can play in helping to meet increased global demands for food’.
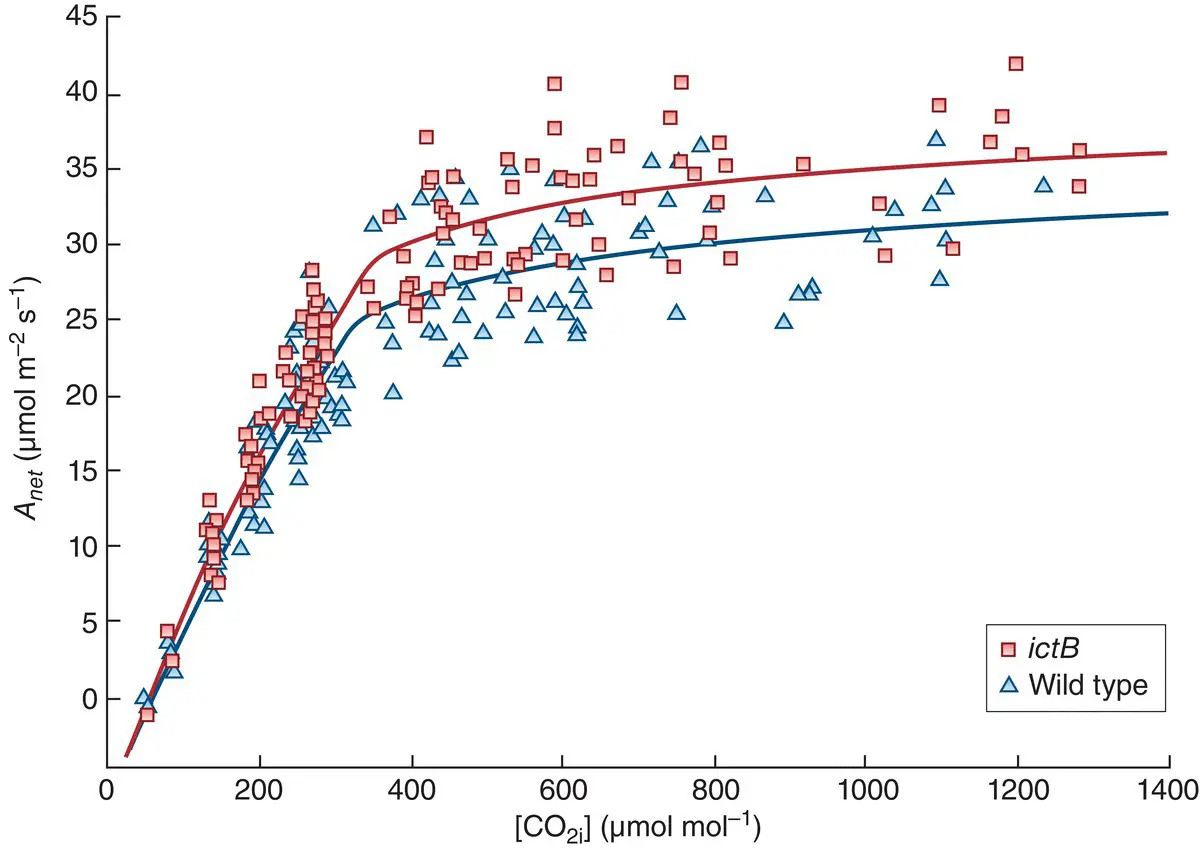
Figure 3.19 Bioengineering of a gene from cyanobacteria into soybean increases its rate of photosynthesis.A comparison of soybean ( Glycine max ) wild type (blue) and a variety genetically engineered to express the cyanobacterial membrane protein ictB (red) in terms of the effect of a plant’s internal CO 2([CO 2i]) concentration on its net rate of photosynthesis ( A net). The fitted lines were non‐rectangular hyperbolas reflecting current understanding of the underlying physiology.
Source : After Hay et al . (2017).
the evolution of C 4and CAM
The evolution of the C 4and CAM pathways, and of CCMs generally (that increase the concentration of CO 2around RuBisCO), has been reviewed by Raven et al . (2008). They describe the very strong evidence that these mechanisms are evolutionarily primitive (the earliest appearing at least 300 million years ago) but also that the C 4and CAM systems must have arisen repeatedly and independently during the evolution of the plant kingdom – the most recent being the appearance of C 4in land plants only 20–30 million years ago. This prolonged coexistence of multiple paths to carbon fixation foreshadows a pattern we will see many more times in later chapters – of coexisting species utilising the same resources but in different ways.
3.4.2 The response of plants to changing atmospheric concentrations of CO 2
Интервал:
Закладка:
Похожие книги на «Ecology»
Представляем Вашему вниманию похожие книги на «Ecology» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Ecology» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.