Fermentation Processes - Emerging and Conventional Technologies
Здесь есть возможность читать онлайн «Fermentation Processes - Emerging and Conventional Technologies» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Fermentation Processes: Emerging and Conventional Technologies
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Fermentation Processes: Emerging and Conventional Technologies: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Fermentation Processes: Emerging and Conventional Technologies»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
The text begins by covering the conventional fermentation process, discussing cell division and growth kinetics, current technologies and developments in industrial fermentation processes, the parameters and modes of fermentation, various culture media, and the impact of culture conditions on fermentation processes. Subsequent chapters provide in-depth examination of the use of emerging technologies—such as pulsed electric fields, ultrasound, high-hydrostatic pressure, and microwave irradiation—for biomass fractionation and microbial stimulation. This authoritative resource:
Explores emerging technologies that shorten fermentation time, accelerate substrate consumption, and increase microbial biomass Describes enhancing fermentation at conventional conditions by changing oxygenation, agitation, temperature, and other medium conditions Highlights the advantages of new technologies, such as reduced energy consumption and increased efficiency Discusses the integration and implementation of conventional and emerging technologies to meet consumer and industry demand Offers perspectives on the future direction of fermentation technologies and applications
is ideal for microbiologists and bioprocess technologists in need of an up-to-date overview of the subject, and for instructors and students in courses such as bioprocess technology, microbiology, new product development, fermentation, food processing, biotechnology, and bioprocess engineering.
Fermentation Processes: Emerging and Conventional Technologies — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Fermentation Processes: Emerging and Conventional Technologies», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
1.2.4 Fermentation
Some microbial species can obtain their energy in the absence of O 2through the catabolic pathway of fermentation. The only difference compared to respiration is in the final electron acceptor (Angelidaki et al. 2011; Dunford 2012; Madigan et al. 2015). In this case, ATP is produced without the Krebs cycle or an electron transport chain involved. This metabolic pathway does not require O 2, because ATP comes exclusively from glycolysis, and the last electron acceptor is an organic molecule such as pyruvic acid (or a derived molecule). Figure 1.4summarizes the metabolic pathway of fermentation.
As shown in Figure 1.4, glucose is oxidized during glycolysis to form two molecules of pyruvate. The electrons and protons released during this pathway are captured by the NAD +to be reduced to NADH + H +. As shown above, two molecules of ATP are produced during glycolysis. To regenerate the NAD +, the NADH + H +must be reoxidized; otherwise, the oxidation of glucose will stop and glycolysis too. During this oxidation, electrons and protons are directly transferred to pyruvate or one of its derivatives. The reduction of these final electron acceptors results in the formation of many different compounds, which provide a great variety of types of fermentation. At the same time, the NAD +is regenerated and can engage in another round of glycolysis. The goal is to provide an uninterrupted supply of NAD +, which allows uninterrupted oxidation of glucose.
During fermentation, all ATP is produced solely by glycolysis, which implies a much lower energy yield compared to aerobic respiration (2 mol of ATP against 38 in prokaryotes). Considering that glucose oxidation is partial, a large part of the energy originally contained in glucose remains stored in the chemical bonds of the final fermentation product (e.g. ethanol, lactic acid, etc.). Fermentation microorganisms must, therefore, compensate for this shortfall by the oxidation of a larger quantity of substrate.
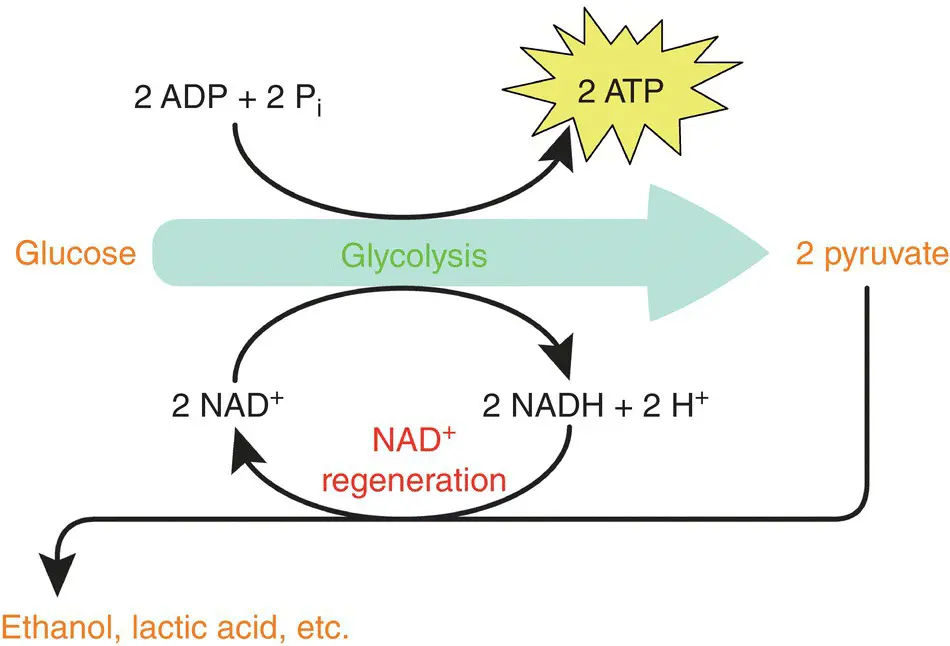
Figure 1.4 Schematic representation of fermentation and energy generation.
Different microorganisms can metabolize organic substrates (e.g. monosaccharides, amino acids, glycerol, etc.) (Angelidaki et al. 2011) to produce organic products such as acids, alcohols, and gases (Wilkins and Atiyeh 2012). This transformation occurs when all essential conditions and factors (e.g. temperature, pH, sugar concentration, culture medium, dissolved O 2, and other micronutrients) required to the growth of microorganisms are provided (Smith 2009). Besides, the microorganism used must be viable, genetically stable, and able to resist several factors including the high concentrations of substrate, salt, and product, and sometimes to the presence of inhibitors, especially when using an industrial by‐product, or a lignocellulosic hydrolyzate.
Under anaerobic conditions, pyruvate is fermented to a wide range of fermentation products; most of them are of industrial importance ( Figure 1.5). In general, microorganisms can be grouped into two classes according to the number of compounds that will be produced during fermentation: homofermentaries and heterofermentaries. Homofermentative microorganisms use a fermentation pathway that leads to the production of a single compound. For example, some lactic acid bacteria can oxidize glucose to produce only lactic acid. Heterofermentative microorganisms, on the other hand, use a fermentation pathway that generates several compounds after the oxidation of the substrate. For example, the bacterium E. coli can produce several types of organic acids during the fermentation of glucose. The following are among the most occurring industrial fermentative products: ethanol, lactic acid, propionic acid, butyric acid, and acetone (Eş et al. 2017; Lin et al. 2014; Navarrete‐Bolaños et al. 2013; Ruijschop et al. 2008).
1.3 Microorganisms Used in Fermentation Processes
Fermentation can take place under anaerobic or aerobic conditions with the help of microorganisms (Gänzle 2015; Ghosh et al. 2015; Kutyna et al. 2012). The most used microorganisms in bioprocesses are bacteria and fungi (yeasts and molds).
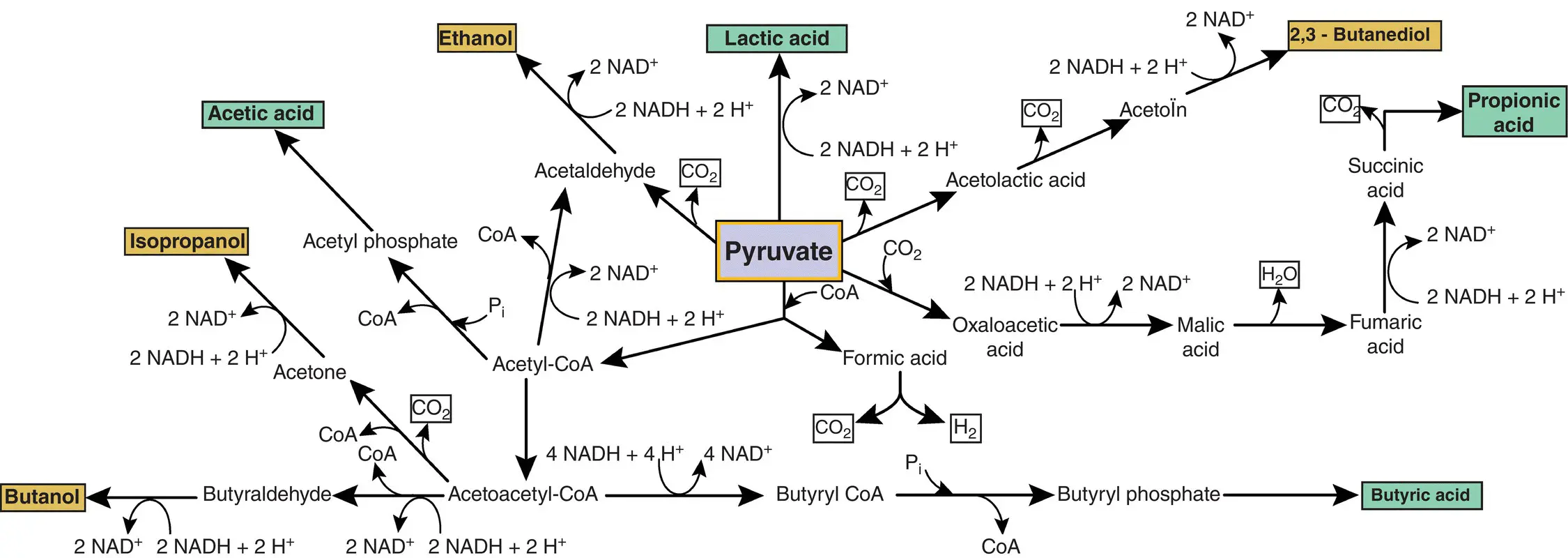
Figure 1.5 Main terminal reactions of catabolic fermentations using pyruvate.
1.3.1 Bacteria
Bacteria are prokaryotes ( Figure 1.6a) and include two distinct categories: archaebacteria or archaea (“ancient” bacteria) and eubacteria (“true” bacteria). The group containing almost all the species used in industrial bioprocesses is eubacteria (Waites et al. 2001) and will, therefore, be detailed below.
Eubacteria group could be divided into 12 subgroups: (i) Proteobacteria; (ii) Gram‐positive eubacteria; (iii) Cyanobacteria; (iv) Chlamydia; (v) Planctomyces and Pirella; (vi) Bacteroides and Flavobacteria; (vii) Green sulfur bacteria; (viii) Spirochetes and relatives; (ix) Deinococci, radioresistant micrococci, and relatives; (x) Green nonsulfur bacteria and anaerobic phototrophs; (xi) Thermotoga and Thermosulfobacteria; and (xii) Aquifex. Only two of them are of industrial interest: the proteobacteria and the Gram‐positive eubacteria.
1.3.1.1 The Proteobacteria
This subgroup constitutes a major kingdom of Gram‐negative bacteria that include purple photosynthetic and nonphotosynthetic bacteria such as the Enterobacteriaceae (e.g. E. coli ), along with Pseudomonas , Hyphomicrobium , Thiobacillus , Nitrobacter , and Vibrio (Waites et al. 2001).
1.3.1.2 The Gram‐Positive Eubacteria
This subgroup could be divided into two major subdivisions. The first one corresponds to the bacteria having low guanine (G) and cytosine (C) base pair content in their DNA and includes Bacillus , Staphylococcus , Leuconostoc , Streptococcus , Clostridium , Lactobacillus , and Mycoplasma . The second subdivision corresponds to the bacteria having high guanine (G) and cytosine (C) base pair content in their DNA and includes the actinomycetes (filamentous bacteria, e.g. Streptomyces ), Mycobacterium , Micrococcus , and Corynebacterium .
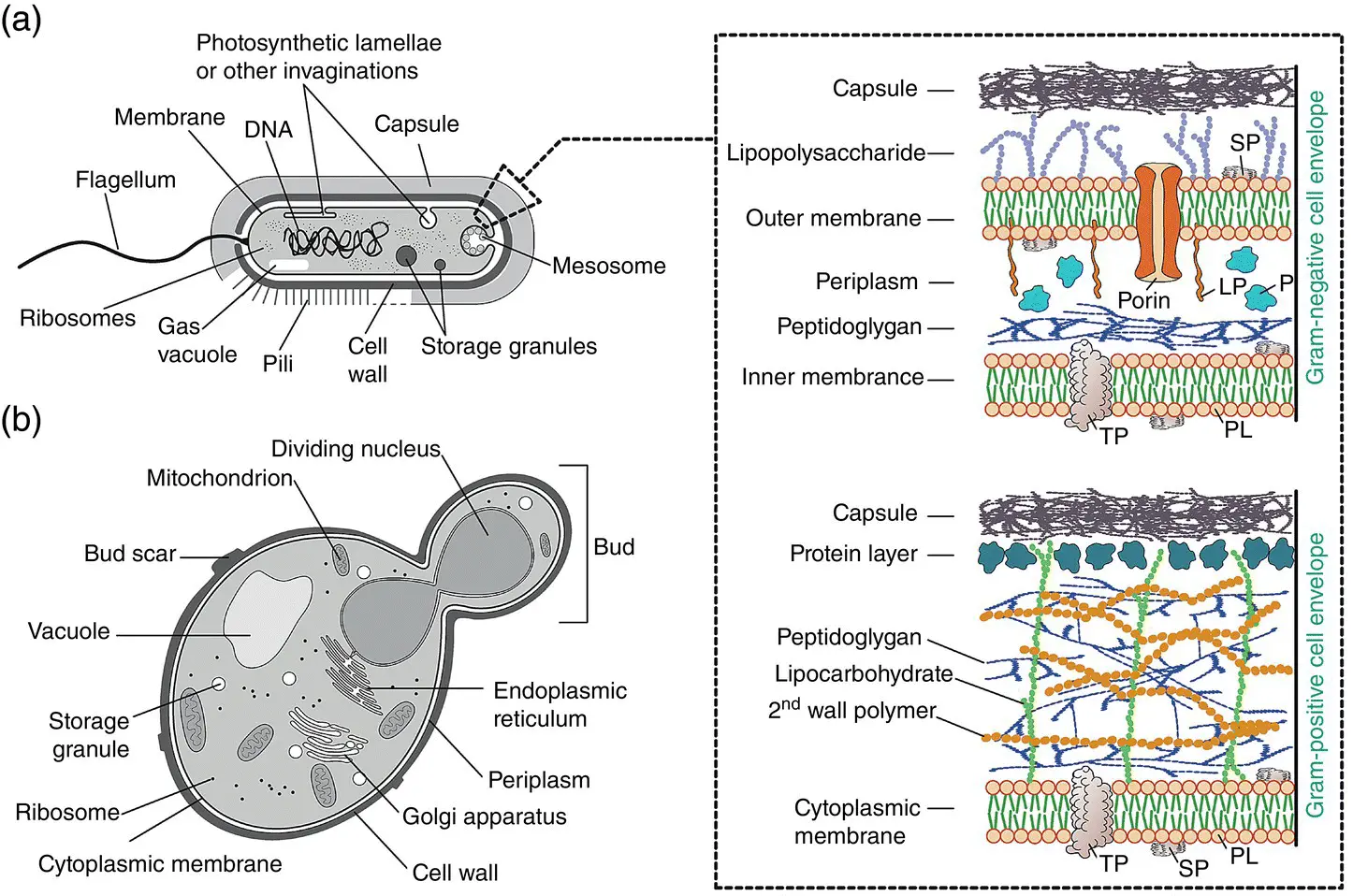
Figure 1.6 Schematic representation of a prokaryotic cell (a) and a budding yeast cell (b). Insert represents a schematic structure of typical Gram‐negative and Gram‐positive cell envelopes. LPS = lipopolysaccharide; LP = lipoprotein; P = protein; PL = phospholipid; SP = surface protein; TP = transmembrane proteins.
The physiological difference between Gram‐negative and Gram‐positive bacteria is mainly in the composition of their cell envelopes (see insert in Figure 1.6).
1.3.2 Fungi
Fungi are eukaryotic microorganisms that could be divided into filamentous hyphae (also called molds ) and unicellular fungi (also called yeasts). A relatively low number of filamentous fungi are used at industrial scale in bioprocesses (e.g. Acremonium , Agaricus , Aureobasidium , Aspergillus , Claviceps , Coniothyrium , Curvularia , Cylindrocarpon , Fusarium , Lentinus , Mortierella , Mucor , Paecilomyces , Penicillium , Rhizomucor , Rhizopus , Sclerotium , Trametes , Trichoderma , and Trichosporon ). Filamentous fungi are chemoheterotrophs and nonphotosynthetic. Most of them secrete a wide variety of hydrolytic enzymes (e.g. cellulose, amylase, xylanase, etc.) (de Souza and de Oliveira Magalhães 2010; Payne et al. 2015; Polizeli et al. 2005) that can degrade different polymers (e.g. lignocellulosic materials) into smaller molecules (e.g. monosaccharides, disaccharides, etc.), which are easily absorbed and metabolized. Filamentous fungi germinate from either individual spore or a fragment of hyphae when exposed to suitable environmental conditions (e.g. pH, temperature, etc.). The length of hyphae can increase rapidly at rates reaching several μm/min (Waites et al. 2001). Further reading about the filamentous fungi can be found in Quintanilla et al. (2015).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Fermentation Processes: Emerging and Conventional Technologies»
Представляем Вашему вниманию похожие книги на «Fermentation Processes: Emerging and Conventional Technologies» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Fermentation Processes: Emerging and Conventional Technologies» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.