Pascal Ribéreau-Gayon - Handbook of Enology - Volume 1
Здесь есть возможность читать онлайн «Pascal Ribéreau-Gayon - Handbook of Enology - Volume 1» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Handbook of Enology: Volume 1
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Handbook of Enology: Volume 1: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Handbook of Enology: Volume 1»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
is thus the same. It aims to provide practitioners, winemakers, technicians and enology students with foundational knowledge and the most recent research results. This knowledge can be used to contribute to a better definition of the quality of grapes and wine, a greater understanding of chemical and microbiological parameters, with the aim of ensuring satisfactory fermentations and predicting the evolution of wines, an7thd better mastery of wine stabilization processes. As a result, the purpose of this publication is to guide readers in their thought processes with a view to preserving and optimizing the identity and taste of wine and its aging potential.
This third English edition of
, is an enhanced translation from the 7h French 2017 edition, and is published in print as individual themed volumes and as a two-volume set, describing aspects of winemaking using a detailed, scientific approach. The authors, who are highly-respected enologists, examine winemaking processes, theorizing what constitutes a perfect technique and the proper combination of components necessary to produce a quality vintage. They also illustrate methodologies of common problems, revealing the mechanism behind the disorder, thus enabling a diagnosis and solution.
Volume 1:
The Microbiology of
Wine and Vinifications
Coverage includes: Wine microbiology; Yeasts; Yeast metabolism; The conditions for the development of yeasts; Lactic acid bacteria, their metabolism and their development in wine; Acetic bacteria; The use of sulfur dioxide in the treatment of musts and wines; Products and processes acting in addition to sulfur dioxide; Winemaking; The grape and its maturation; Harvesting and processing of grapes after harvest; Vinification in red and white wine making.
The target audience includes advanced viticulture and enology students, professors and researchers, and practicing grape growers and vintners.
Handbook of Enology: Volume 1 — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Handbook of Enology: Volume 1», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
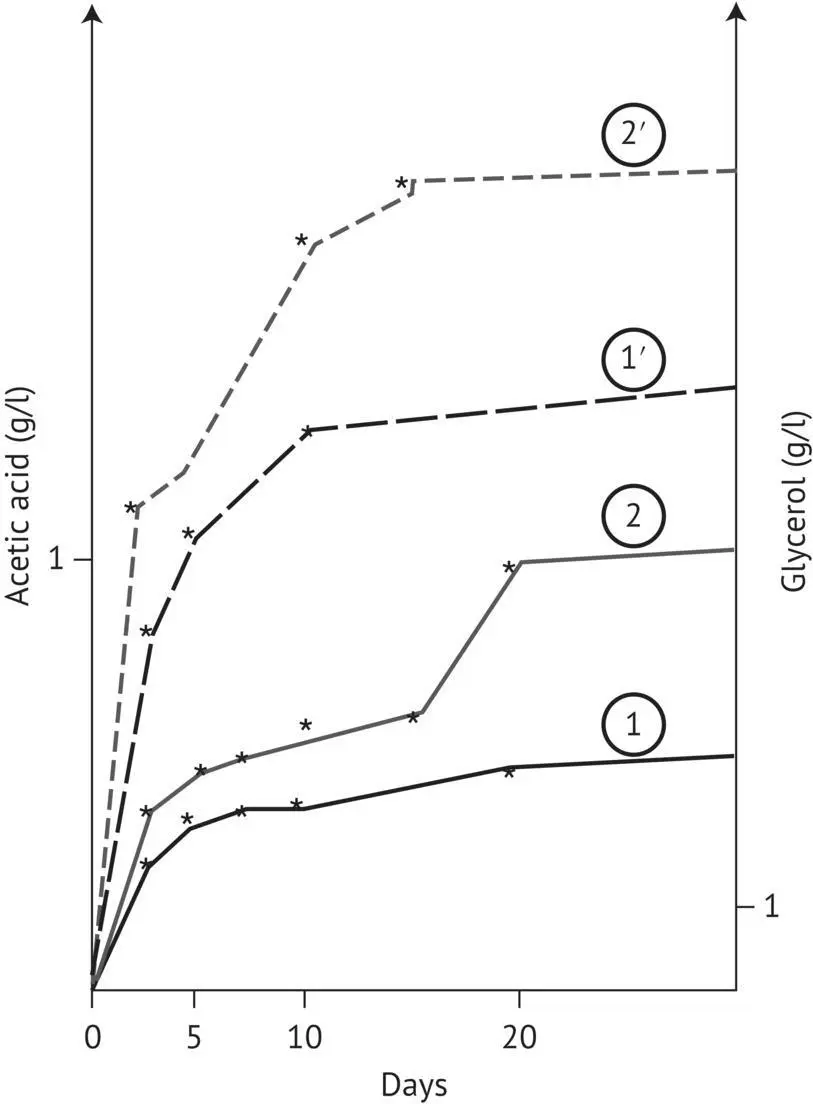
FIGURE 2.15 Effect of an alcohol‐induced precipitate of a botrytized grape must on glycerol and acetic acid formation during the alcoholic fermentation of healthy grape must (Dubourdieu, 1982). 1, Evolution of acetic acid concentration in the control must; 2, evolution of acetic acid concentration in the must supplemented with the freeze‐dried precipitate; 1′, evolution of glycerol concentration in the control must; 2′, evolution of glycerol concentration in the must supplemented with the freeze‐dried precipitate.
Other winemaking factors favor the production of acetic acid by S. cerevisiae : anaerobic conditions, very low pH ( < 3.1) or very high pH (>4), certain amino acid or vitamin deficiencies in the must, and excessively high temperature (25–30°C) during the yeast growth phase. In red winemaking, temperature is the most important factor, especially when the must has a high sugar concentration. In hot climates, the grapes should be cooled when filling the tanks. The temperature should not exceed 20°C at the beginning of fermentation. The same procedure should be followed during thermovinification immediately following the heating of the grapes.
In dry white and rosé winemaking, excessive must clarification can also lead to the excessive production of volatile acidity by yeast. This phenomenon can be particularly pronounced with certain yeast strains. Therefore, must turbidity should be adjusted to the lowest possible level that enables a complete and rapid fermentation (Sections 3.7.3 and 13.5.3). The input of lipids made available to yeasts via solid sediments (grape solids), in particular long‐chain unsaturated fatty acids (C18:1 and C18:2), greatly influences acetic acid production during white and rosé winemaking.
The experiment in Figure 2.16illustrates the important role of lipids in acetic acid metabolism (Delfini and Cervetti, 1992; Alexandre et al ., 1994). The volatile acidity of three wines obtained from the same Sauvignon Blanc must was compared. After filtration but before yeast inoculation, must turbidity was adjusted to 250 Nephelometric Turbidity Units (NTU) by three different methods: reincorporating fresh grape solids (control), adding cellulose powder, and supplementing with a lipid extract (using methanol–chloroform) that contained the same quantity of grape solids adsorbed on the cellulose powder. The volatile acidities of the control wine and the wine that was supplemented with a lipid extract of grape solids before fermentation are identical and perfectly normal. Although the fermentation was normal, the volatile acidity of the wine made from the must supplemented with cellulose (therefore devoid of lipids) was practically twice as high (Lavigne, 1996). Supplementing the medium with lipids appears to favor the penetration of amino acids into the cell, which limits the formation of acetic acid.
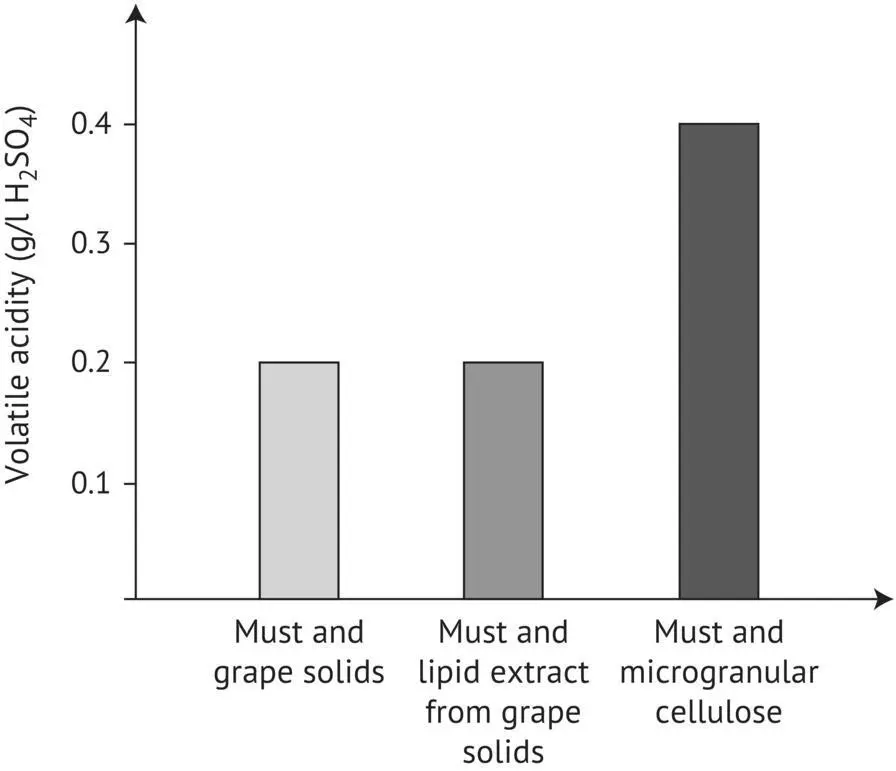
FIGURE 2.16 Effect of the lipid fraction of grape solids on acetic acid production by yeasts during alcoholic fermentation (Lavigne, 1996).
During the alcoholic fermentation of red or slightly clarified white wines, yeasts do not continuously produce acetic acid. The yeast metabolizes a large portion of the acetic acid secreted in must during the fermentation of the first 50–100 g of sugar. It can also assimilate acetic acid added to must at the beginning of alcoholic fermentation. The assimilation mechanisms are not yet clear. Acetic acid appears to be reduced to acetaldehyde, which favors alcoholic fermentation to the detriment of glyceropyruvic fermentation. In fact, the addition of acetic acid to a must lowers glycerol production but increases the formation of acetoin and 2,3‐butanediol. Yeasts seem to use the acetic acid formed at the beginning of alcoholic fermentation (or added to must) via acetyl‐CoA in their lipid synthesis pathways.
Certain winemaking conditions produce abnormally high amounts of acetic acid. Since this acid is not used during the second half of the fermentation, it accumulates until the end of fermentation. When refermenting a tainted wine, yeasts can lower volatile acidity by metabolizing acetic acid. The wine is incorporated into freshly crushed grapes at a proportion of no more than 20–30%. The wine should be sulfited or filtered before incorporation to eliminate bacteria. The volatile acidity of this mixture should not exceed 0.6 g/l expressed as H 2SO 4. The volatile acidity of this newly made wine rarely exceeds 0.3 g/l expressed as H 2SO 4. The concentration of ethyl acetate decreases simultaneously.
2.3.5 Other Secondary Products of the Fermentation of Sugars
Lactic acid is another secondary product of fermentation. It is also derived from pyruvic acid, which is directly reduced by yeast L(+)‐ and D(−)‐lactate dehydrogenases. Under anaerobic conditions (the case in alcoholic fermentation), the yeast synthesizes predominantly D(−)‐lactate dehydrogenase. Yeasts form 200–300 mg of D(−)‐lactic acid per liter and only a few dozen milligrams of L(+)‐lactic acid. The latter is formed essentially at the start of fermentation. By determining the D(−)‐lactic acid concentration in a wine, it can be ascertained whether the origin of acetic acid is yeast or lactic acid bacteria (Section 14.2.3). Wines that have undergone malolactic fermentation can contain several grams per liter of exclusively L(+)‐lactic acid. On the other hand, the lactic acid fermentation of sugars (lactic spoilage) forms D(−)‐lactic acid. When D(−)‐lactic acid concentrations exceed 200–300 mg/l, it is clear that lactic acid bacteria have transformed substrates other than malic acid.
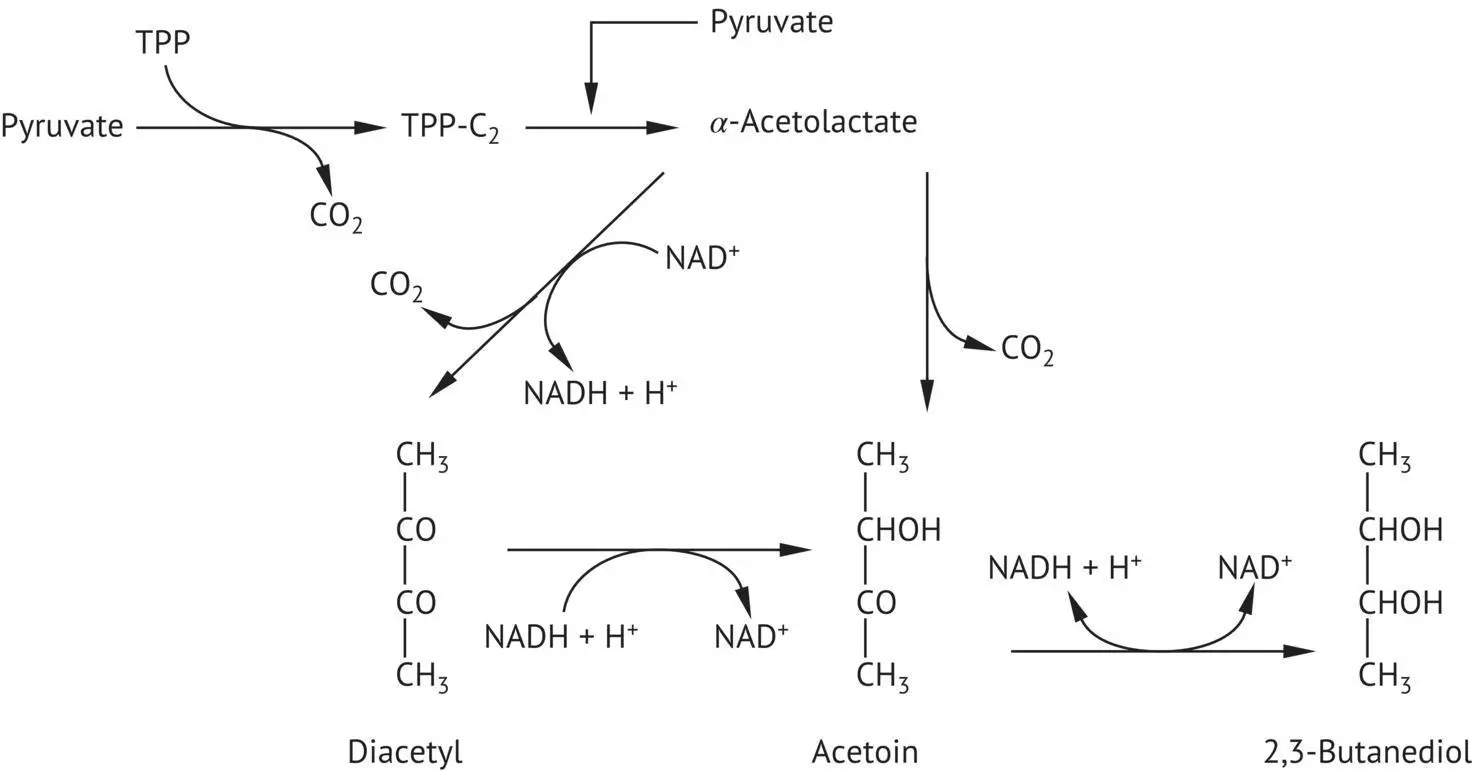
FIGURE 2.17 Acetoin, diacetyl, and 2,3‐butanediol formation by yeasts under anaerobic conditions. TPP, thiamine pyrophosphate; TPP‐C 2, active acetaldehyde.
Yeasts also make use of pyruvic acid to form acetoin, diacetyl, and 2,3‐butanediol ( Figure 2.17). This process begins with the condensation of a pyruvate molecule and a molecule of active acetaldehyde bound to TPP, leading to the formation of α ‐acetolactic acid. The oxidative decarboxylation of α ‐acetolactic acid produces diacetyl. Acetoin is produced by either the non‐oxidative decarboxylation of α ‐acetolactic acid or the reduction of diacetyl. The reduction of acetoin leads to the formation of 2,3‐butanediol; this last reaction is reversible.
From the start of alcoholic fermentation, yeasts produce diacetyl, which is rapidly reduced to acetoin and 2,3‐butanediol. This reduction takes place in the days that follow the end of alcoholic fermentation, when wines are conserved on the yeast biomass (de Revel et al ., 1996). Acetoin and especially diacetyl are strong‐smelling compounds that evoke a buttery aroma. Above a certain concentration, they have a negative effect on wine aroma. However, in wines that have not undergone malolactic fermentation, their concentration is too low (a few milligrams per liter for diacetyl) to have a sensory impact. On the other hand, lactic acid bacteria can degrade citric acid to produce much higher quantities of these carbonyl compounds than yeasts (Section 5.3.2).
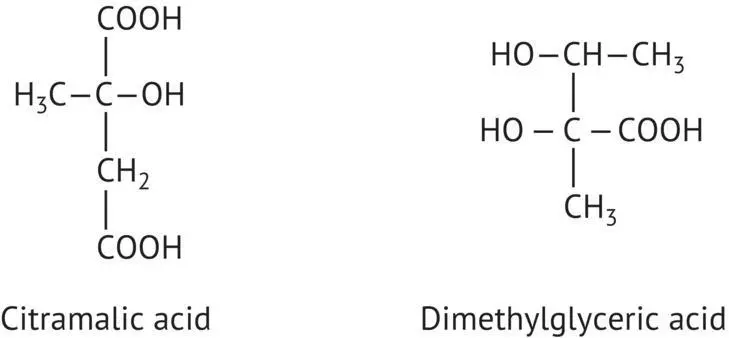
FIGURE 2.18 Citramalic acid and dimethylglyceric acid.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Handbook of Enology: Volume 1»
Представляем Вашему вниманию похожие книги на «Handbook of Enology: Volume 1» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Handbook of Enology: Volume 1» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.