Robert E. Blankenship - Molecular Mechanisms of Photosynthesis
Здесь есть возможность читать онлайн «Robert E. Blankenship - Molecular Mechanisms of Photosynthesis» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Molecular Mechanisms of Photosynthesis
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Molecular Mechanisms of Photosynthesis: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Molecular Mechanisms of Photosynthesis»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Molecular Mechanisms of Photosynthesis — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Molecular Mechanisms of Photosynthesis», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
4.3 Chlorophyll biosynthesis
The chlorophyll biosynthetic pathway contains 17 enzymatic steps, as shown in Fig. 4.6. It begins with the formation of δ ‐aminolevulinic acid (ALA) (Suzuki et al ., 1997; Beale, 1999; Chew and Bryant, 2007; Masuda, 2008; Chen, 2014; Bryant et al ., 2020). Eight molecules of ALA are condensed, eventually forming the symmetric metal‐free porphyrin: protoporphyrin IX. At this point the pathway branches, with one branch leading to heme and the other to chlorophyll, depending on whether Fe or Mg is incorporated. The chlorophyll branch includes additional steps in which the fifth ring is fashioned and a double bond in ring D is reduced to form chlorophyllide, which still lacks the isoprenoid tail. The isoprenoid tail is attached as the last step. Biosynthesis of bacteriochlorophyll a is identical to chlorophyll synthesis through the chlorophyllide a intermediate, but contains two additional steps: the reduction of ring B and the conversion of the vinyl group at C‐3 to an acetyl moiety. The order of the biosynthetic steps and the identities of most of the enzymes that carry out the transformations have been well established for both chlorophyll a and bacteriochlorophyll a synthesis by a combination of biochemical and genetic experiments. The biosynthetic pathway of chlorophylls is shown in Fig. 4.6. The enzymes that catalyze the various steps are identified in the figure legend.
The first few steps in the pathway produce ALA. Most photosynthetic organisms use a unique pathway that involves the ATP‐dependent charging of the amino acid glutamic acid to a glutamyl tRNA, the same reaction that takes place when glutamic acid is incorporated into a growing peptide during protein synthesis. This reaction is one of a very small number of reactions known in biology in which a tRNA molecule is used in a biochemical step other than protein synthesis. Step 2 is the NADPH‐dependent reductive cleavage of the acid to an aldehyde to form L‐glutamic acid 1‐semialdehyde. The final step in ALA synthesis is the transamination rearrangement to form ALA.
Surprisingly, one group of photosynthetic organisms makes ALA by a completely different route. This is the one‐step condensation of the amino acid glycine with succinyl‐CoA, followed by decarboxylation, catalyzed by the enzyme ALA synthase. This enzyme is found in some (but not all) purple photosynthetic bacteria. It is also found in the mitochondrion of all eukaryotic cells and is the pathway used by mitochondria to make ALA for heme biosynthesis.
The next series of biosynthetic steps lead to protoporphyrin IX. Step 4 is the condensation of two molecules of ALA to form the cyclic compound porphobilinogen, which represents the pyrrole monomer. Step 5 is the further condensation of four molecules of porphobilinogen to form the open‐chain tetrapyrrole hydroxymethylbilane. Steps 6–8 involve ring closure and successive oxidative decarboxylation steps. Note the remarkable “flipping” of pyrrole ring D during step 6, so that the positions of the acetate and propionate substituents are interchanged. This proceeds by way of a spiro intermediate at C‐16.
The final step in this phase of the biosynthetic pathway is the oxidation of protoporphyrinogen IX to protoporphyrin IX, step 9 in the sequence shown in Fig. 4.6. This is an important step, because the former compound looks superficially like a porphyrin, but is not fully conjugated. The pyrrole rings are essentially independent of each other, and the compound is colorless. The product, protoporphyrin IX, is a fully conjugated porphyrin and is highly colored. Excited protoporphyrin IX reacts readily with molecular oxygen to form the highly damaging species singlet oxygen. The system is highly regulated so that high concentrations of free protoporphyrin IX and other photosensitive intermediates do not build up.
Protoporphyrin IX is metallated by insertion of Mg at step 10. The heme biosynthesis pathway branches at this point (Bryant et al ., 2020). Those molecules destined to become heme have Fe inserted instead of Mg. The next phase of the biosynthesis of chlorophylls involves the construction of the isocyclic ring E by cyclizing the propionic acid attached to C‐13. This reaction proceeds by first esterifying the carboxylic acid moiety and then undergoing a stereospecific oxidative cyclase reaction, steps 12–14. The intermediate at this step, divinyl protochlorophyllide, is then acted on by two separate enzymes on opposite sides of the ring. The two enzymes are not sensitive to whether the other change has been made in the substrate, so the pathway branches, and a given molecule can first have step 15 and then step 16 take place, or vice versa. Step 15 is the reduction of the vinyl group at C‐8 to an ethyl group, and step 16 is the reduction of ring D.
The reduction of ring D is one of the most interesting steps in the biosynthesis of chlorophylls. In all oxygenic photosynthetic organisms, a light‐driven enzyme, protochlorophyllide reductase, (often called POR), carries out this step. This enzyme uses NADPH as a source of reductant but also has an absolute requirement for light, making it one of a very small number of light‐driven enzymes known in all of biology (another is the DNA repair enzyme photolyase). The structure of the light‐driven POR from a cyanobacterium has been determined (Dong et al ., 2020). In anoxygenic photosynthetic bacteria, an entirely different enzyme complex that is not light‐driven carries out this reduction. In this case, the reductant is reduced ferredoxin instead of NADPH. All oxygenic organisms except higher plants have both the light‐dependent and the light‐independent enzymes. The evolutionary implications of this pattern are discussed in Chapter 12.
The final step in chlorophyll a biosynthesis is the attachment of the tail, catalyzed by the enzyme chlorophyll synthase and using phytol pyrophosphate as the substrate. Phytol pyrophosphate is made by reduction of the unsaturated polyisoprenoid compound geranylgeranyl pyrophosphate. Depending on the state of growth of the organism, the reduction can take place either before or after attachment of the tail to the macrocycle.
The same basic pathway is used for the synthesis of all other chlorophylls and bacteriochlorophylls. Most of the reactions are the same, except that some steps are O 2‐dependent in aerobes but use a different enzyme and oxidant in anaerobes (Ouchane et al ., 2004; Raymond and Blankenship, 2004). The major differences are found at the end of the pathway. Bacteriochlorophyll a is made from chlorophyllide a using two additional steps. The first is the conversion of the C‐3 vinyl group to an acetyl group. The second is the reduction of pyrrole ring B by an enzyme complex similar to the light‐independent enzyme that reduces ring D, as discussed in the previous paragraph.
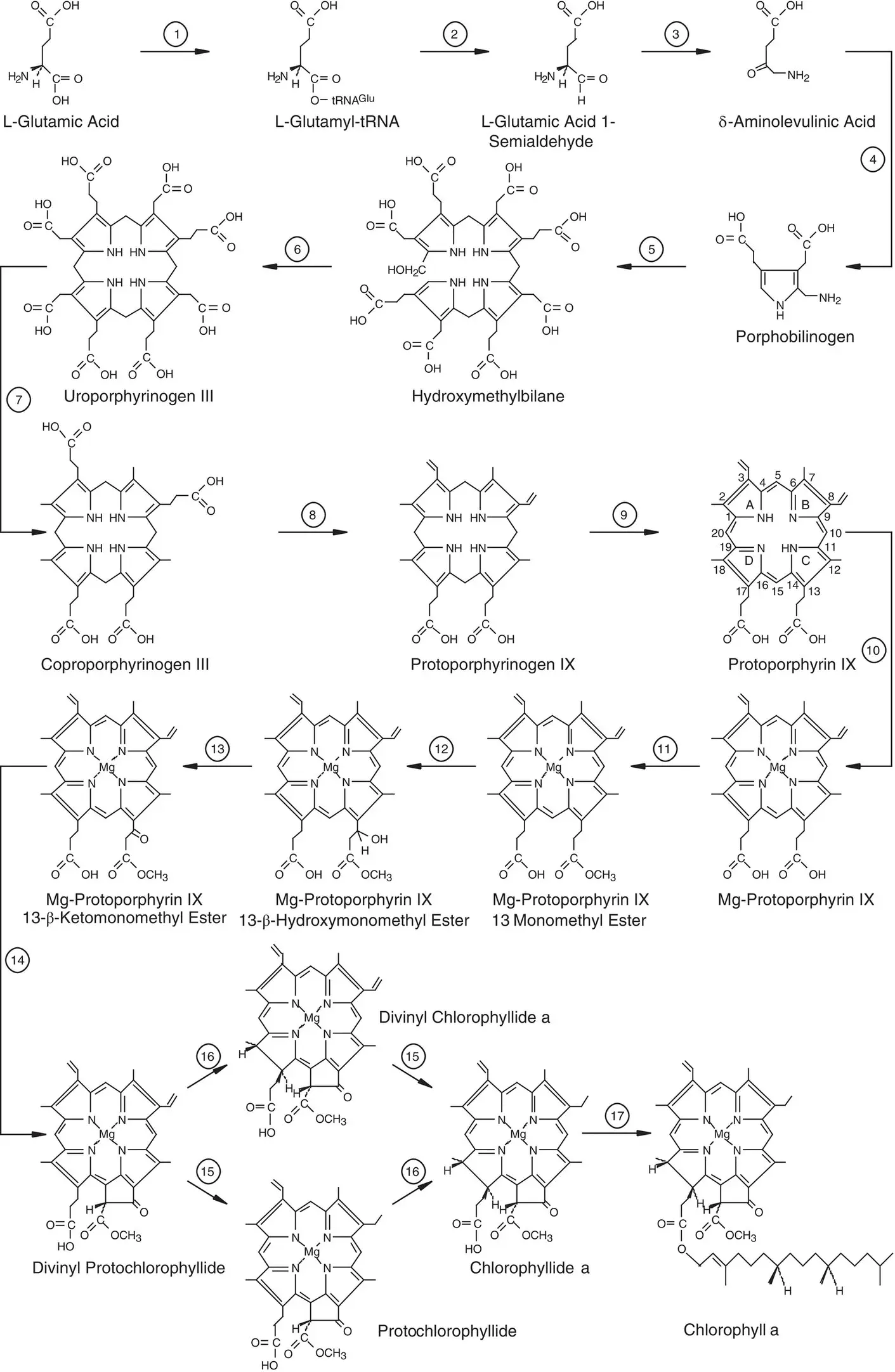
Figure 4.6 Outline of chlorophyll a biosynthesis from glutamate. The enzymes that catalyze the individual numbered reactions are (1) glutamyl‐tRNA synthetase; (2) glutamyl‐tRNA reductase; (3) glutamate 1‐semialdehyde aminotransferase; (4) porphobilinogen synthase; (5) hydroxymethylbilane synthase; (6) uroporphyrinogen III synthase; (7) uroporphyrinogen III decarboxylase; (8) coproporphyrinogen III oxidative decarboxylase; (9) protoporphyrinogen IX oxidase; (10) protoporphyrin IX Mg‐chelatase;(11) S‐adenosyl‐L‐methionine:Mg‐protoporphyrin IX methyl‐transferase; (12)–(14) Mg‐protoporphyrin IX monomethyl ester oxidative cyclase; (15) divinyl (proto)chlorophyllide 4‐vinyl reductase;(16) light‐dependent NADPH:protochlorophyllide oxidoreductase or light‐independent protochlorophyllide reductase; (17) chlorophyll synthase. The IUPAC numbering for the tetrapyrrole peripheral substituent positions is shown for protoporphyrin IX.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Molecular Mechanisms of Photosynthesis»
Представляем Вашему вниманию похожие книги на «Molecular Mechanisms of Photosynthesis» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Molecular Mechanisms of Photosynthesis» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.