Nitric Oxide in Plants
Здесь есть возможность читать онлайн «Nitric Oxide in Plants» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Nitric Oxide in Plants
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Nitric Oxide in Plants: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Nitric Oxide in Plants»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Examines the beneficial roles of nitric oxide in growth and stress tolerance regulation through its involvement in tolerance mechanisms Nitric Oxide in Plants: A Molecule with Dual Roles
Nitric Oxide in Plants: A Molecule with Dual Roles
Nitric Oxide in Plants — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Nitric Oxide in Plants», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
NO production, along with H 2O 2and O 2, was observed to be associated with elicitor-elicited mortality in Mexican cypress. The co-accumulation of ROS and NO, as well as the interplay between H 2O 2, O 2, and NO, mediated the death response. The use of protein scavengers/inhibitors was used to investigate the role of NO and O 2in death. NO and H 2O 2reciprocally promoted each other’s assembly, whereas NO and O 2reciprocally suppressed each other’s production. The interaction between NO and O 2, but not between NO and H 2O 2, triggered PCD via peroxynitrite (ONOO −) (Neill et al. 2002; Zhao 2005). The stress response in bacterially triggered PCD in soybean and Arabidopsis concerned reactions to NO and H 2O 2, with interactions between NO and H 2O 2being synergistic and additive in different ways, which is consistent with previous findings. After exposing tobacco leaves to a high intensity of stress, NO-mediated death was seen in enzyme-deficient (CAT1AS) plants but not in wild types, revealing the interaction of NO with H 2O 2throughout the NO-mediated death response (Zago et al. 2006).
Plants protect themselves from microorganisms by triggering elaborate defenses. Similarly, it was recently demonstrated that NO works synergistically with ROS to prolong host mortality in soybean suspension cells, and that NOS inhibitors impair the hypersensitized resistance response in Arabidopsis and tobacco (Delledonne et al. 1998). It was discovered that the host plant has intracellular defense mechanisms to combat the oxidative stress caused by pathogens. Increased activity of the catalyst inhibitor system, total phenol content, and phenyl ammonia lyase activity slowed oxidative burst, which was presumably mediated by excessive chemical group content and hence involved gas signaling; this explained the protection against pathogen stress (Dwivedi et al. 2016). Following that, extensive studies revealed that NO could influence the expression of genes encoding a variety of plant effector and restriction proteins (Polverari et al. 2003; Huang et al. 2004; Parani et al. 2004; Zago et al. 2006). Furthermore, recent research on catalase-deficient tobacco plants discovered a small number of genes that are particularly controlled by NO or H 2O 2(Zago et al. 2006). Interestingly, most of the notable genes were affected by both NO and H 2O 2, showing a significant overlap in these two separate chemical process signaling agents (Figure 1.2).
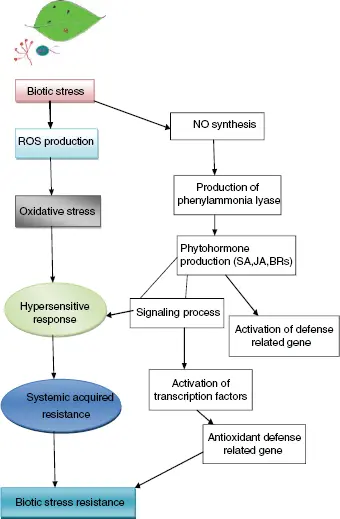
Figure 1.2 Schematic representation of NO signaling against biotic stress in plants.
1.3.1 Interaction of NO with Other Molecules to Confer Biotic Stress Responses in Plants
Plants defend themselves against microorganism attack by activating elaborate defenses. Similarly, it has been recently demonstrated first that NO acts synergistically with ROS to extend host death in soybean suspension cells, and second that NOS inhibitors compromise the hypersensitized resistance response in Arabidopsis and tobacco (Delledonne et al. 1998). NO and ROS induce SA synthesis, and elevated SA may subsequently result in improved NO and ROS levels. SA additionally induces expression of genes that may be NO targets/sensors (e.g. pathogen-induced oxido-reductase [PIOX]). What is more, SA may enhance the results of NO activity through interaction with many NO X-regulated enzymes (e.g. catalase and aconitase). On the other hand, SA, which is a very important antioxidant in mammals, may counter the effects of NO. As an example, NO blocks respiration via inhibition of cytochrome enzyme, while SA induces production of various NO-resistant enzymes and sensors/targets in plants (e.g. pathogen-induced oxygenase, PIOX). NO and SA have an effect on common targets (e.g. catalase, aconitase). SA counteracts NO action (e.g. NO inhibits cytochrome enzyme, whereas SA induces a different oxidase). The induction of necrobiosis and/or defense factor activation occurs via the action of NO, SA, and ROS. NO is not only thought to perform throughout the event of hypersensitive cell death but is also needed in the establishment of disease resistance. Melatonin also enhances pathogen resistance in plants by increasing the levels of SA and ethylene. Lee et al. (2015) reported that decreased level of melatonin content in Arabidopsis mutant lowers the SA content thereby reducing resistance to pathogen infection. It also stimulates elevation of endogenous NO levels and increased expression of defense-related genes, along with disease resistance against Pst DC3000 infection.
Melatonin confers plants with innate immunity via the NO- and H 2O 2-dependent pathway by stimulating mitogen-activated protein kinase kinase kinase 3 (MAPKKK3) and oxidative signal-inducible1 (OXI1). Melatonin stimulates accumulation of NO and SA and improved tomato TMV resistance (Zhao et al. 2019).
Interaction between NO and glutathione is an important component to the NPR1-dependent defense signaling pathway in A. thaliana (Kovacs et al. 2015). Cao et al. (2019) found that brassinosteroids aid in stimulating the susceptibility of maize to maize chlorotic mottle virus infection in a NO-dependent manner. Nitric oxide further acts as a downstream signaling molecule in brassinosteroid-mediated virus susceptibility to maize chlorotic mottle virus in maize. The brassinosteroids provide systemic resistance against viruses along with H 2O 2and NO in Nicotiana benthamiana (Deng et al. 2016). However, several studies have demonstrated the integrative role of brassinosteroids with NO in response to pathogen attack in various species (Hayat et al. 2010; Shi et al. 2015; Zou et al. 2018; Kohli et al. 2019).
1.4 Nitric Oxide’s Role in Abiotic Stress
Existing climatic aberrations cause plants to be exposed to a variety of abiotic stresses such as drought, temperature increases, salinity, and oxidative damage. Because plants are sessile, they must deal with changing environmental conditions throughout their lives. Though plants develop various strategies to cope with adverse conditions, plant scientists and researchers are discovering new methods to learn about plants in stressful environments. In the presence of salt, substantial metals, drought, and oxidative stress, NO plays a critical role in abiotic stress management (Zhao et al. 2001, 2004; Uhida et al. 2002; Kopyra and Gwóźdź 2003). Abiotic stressors are a key stumbling block to agricultural growth while also jeopardizing food security (Figure 1.3). They also jointly influence the metabolism of plants and the defense of inhibitors. Recent NO studies show that they play a serious role in plant growth and mitigate environmental stress (Aroca et al. 2015; Corpas et al. 2019). Drought tolerance of cut leaves and seedlings of wheat was improved by exogenous NO application (Garcia-Mata et al. 2001). NO also mediates drought tolerance by the activation of numerous enzymes and plant metabolism inhibitors (Filippou et al. 2014; Shi et al. 2014). In many species, NO mediates stomatal rotation and defense methods in water stress (Greco et al. 2012; Garcia-Mata and Lamattina 2013; Chen et al. 2016). ROS and NO work synergistically to address the stress in water that causes ABA synthesis in wheat roots (Zhao et al. 2001). As proof of this, ABA accumulation under stress was prevented by the administration of ROS scavengers. The accumulation of NO tested jointly is needed in broad bean throughout ABA-induced stomata closure (Garcia-Mata and Lamattina 2002). Heat treatment of alfalfa cells, for example, has led to an increase in NO synthesis, while the application of exogenous NO improved cold tolerance in tomato, wheat, and maize (Neill et al. 2003). The observed effects were most likely associated with NO’s antioxidative action, which amplifies the negative effects caused by the intensification of peroxidative metabolism in high temperatures (Neill et al. 2002).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Nitric Oxide in Plants»
Представляем Вашему вниманию похожие книги на «Nitric Oxide in Plants» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Nitric Oxide in Plants» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.