Александр Марков - Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества
Здесь есть возможность читать онлайн «Александр Марков - Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 2019, ISBN: 2019, Издательство: АСТ, Corpus, Жанр: Биология, sci_popular, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.
- Название:Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества
- Автор:
- Издательство:АСТ, Corpus
- Жанр:
- Год:2019
- Город:Москва
- ISBN:978-5-17-114115-8
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Исследование № 18
Бессмысленное усложнение
Данное исследование, на наш взгляд, имеет большое мировоззренческое значение. Оно наглядно показывает, как в ходе эволюции сложное может развиться из простого совершенно случайно и без всякой пользы. Организм усложняется, не получая от этого никакой выгоды: эффективность выполнения всех функций остается на прежнем уровне. Этот пример, добавляя конкретики описанным выше оценкам последствий генных дупликаций (см. Исследование № 17), еще раз подчеркивает «недальновидность» естественного отбора, его работу только «здесь и сейчас». С помощью генно-инженерных экспериментов американские биологи расшифровали последовательность событий, в результате которых у предков пекарских дрожжей ( Saccharomyces cerevisiae ) усложнилась одна из регуляторных систем. Предковый ген удвоился, и в каждой из двух копий стали накапливаться свои мутации. В итоге каждая копия утратила ту или иную часть исходных функций. Функции, утраченные каждой из копий, были разными, благодаря чему копии перестали быть избыточными — теперь оба гена, по-разному подпорченные мутациями, стали жизненно необходимы организму. Дальнейшая специализация двух генов подстегивалась тем, что поначалу они конкурировали, мешая друг другу работать. Минимизация конкуренции потребовала закрепления дополнительных мутаций. В конце концов система усложнилась (два специализированных гена вместо одного многофункционального), хотя сами дрожжи ничего от этого не выиграли. Усложнение стало побочным эффектом цепочки отчасти случайных, отчасти закономерных событий, начало которым положило случайное удвоение гена.


Данное исследование было выполнено биологами из Калифорнийского университета в Сан-Франциско (США) в 2013 году ( Baker et al., 2013). Работа пролила свет на процесс появления эволюционных новшеств в результате генных дупликаций — на два важных его аспекта.
Во-первых, она показала, что генные дупликации могут чисто автоматически, через ряд взаимообусловленных промежуточных шагов, приводить к усложнению регуляторных генных сетей. Это происходит даже в том случае, если в подобном усложнении нет необходимости и организм мог бы продолжать с тем же успехом обходиться простой регуляторной сетью.
Во-вторых, работа показала, что копиям удвоившегося гена (паралогам) не всегда удается легко и быстро разделить функции, особенно если ген включен в сложную сеть межмолекулярных взаимодействий. В этом случае копии, едва начав делить функции, начинают конкурировать и мешать друг другу работать. Для устранения этой проблемы требуется закрепление дополнительных мутаций, что способствует окончательному превращению паралогов в два специализированных белка со строгим разделением обязанностей.
Авторы изучили последствия дупликации гена Mcm 1 у дрожжей. Этот ген есть у всех грибов. Он кодирует важный регуляторный белок, который, объединяясь в комплексы с другими регуляторными белками (кофакторами), присоединяется к ДНК, чтобы активировать близлежащие гены (рис. 18.1). Mcm 1 в целом консервативен, хотя и имеет некоторые различия у разных видов.
У некоторых дрожжей, например Kluyveromyces lactis , гены метаболизма аргинина (ARG) регулируются комплексом из двух белковых молекул Mcm1 (такие комплексы называют гомодимерами ), который, в свою очередь, соединяется с кофактором Arg81 (рис. 18.1, а ). У пекарских дрожжей, Saccharomyces cerevisiae, и их ближайшей родни белковый комплекс, регулирующий работу генов ARG, устроен сложнее. Он включает не два, а три разных белка. Вместо гомодимера из двух молекул Mcm1 пекарские дрожжи используют гетеродимер — соединенные молекулы белков Mcm1 и Arg80 (рис. 18.1, б ), и уже этот гетеродимер соединяется с Arg81.
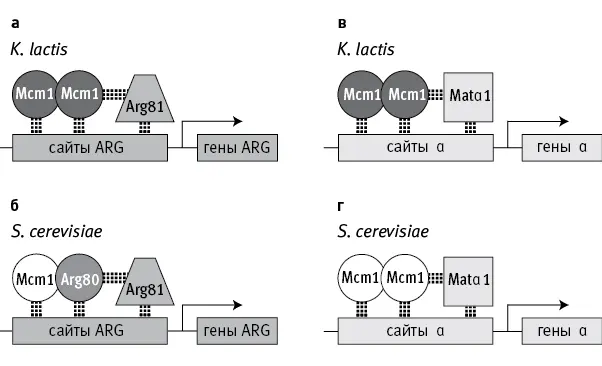
рис. 18.1.Схема регуляции работы генов, связанных с метаболизмом аргинина (гены ARG) и с половым размножением (гены α), у дрожжей Kluyveromyces lactis и Saccharomyces cerevisiae . Комплексы белков-регуляторов присоединяются к регуляторным участкам ДНК (сайты ARG, сайты α) возле контролируемых генов. У K. lactis обе группы генов (ARG и α) регулируются гомодимерами — комплексами из двух молекул белка Mcm1 ( а, в ). У S. cerevisiae гены α регулируются такими же гомодимерами ( г ), а для регуляции генов ARG используется гетеродимер, состоящий из Mcm1 и дополнительного регуляторного белка Arg80, которого нет у K. lactis ( б ). Ген Arg80 появился у предков S. cerevisiae в результате дупликации исходного гена Mcm1 и последующего разделения функций. Arg81 и Matα1 — кофакторы. По рисунку из Baker et al., 2013 .
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества»
Представляем Вашему вниманию похожие книги на «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.










Обсуждение, отзывы о книге «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.