Владимир Бердников - Эволюция и прогресс
Здесь есть возможность читать онлайн «Владимир Бердников - Эволюция и прогресс» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Новосибирск, Год выпуска: 1991, ISBN: 1991, Издательство: Наука, Жанр: Биология, sci_popular, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Эволюция и прогресс
- Автор:
- Издательство:Наука
- Жанр:
- Год:1991
- Город:Новосибирск
- ISBN:5-02-030062-4
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Эволюция и прогресс: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Эволюция и прогресс»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Книга предназначена для биологов широкого профиля, а также всех интересующихся вопросами эволюции живых существ.
Эволюция и прогресс — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Эволюция и прогресс», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
В данной ситуации мутации, повышающие мощность лимитирующей структуры, очевидно становятся благоприятными, так как они повышают уровень относительной приспособленности их носителей. Поэтому коэффициент отбора ( s ) в пользу этих мутаций должен быть близок к относительному приросту мощности лимитирующей структуры, во всяком случае, он должен быть ему пропорционален, т. е.
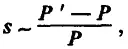 (5.3)
(5.3)
где Р ' — мощность структуры у особей — носителей благоприятной мутации; Р — мощность той же структуры у средней особи популяции. Фактически это означает, что фиксация каждой благоприятной мутации увеличивает мощность лимитирующей структуры в некоторое число раз (на какой-то процент). При продолжающемся ухудшении среды продолжается и процесс накопления благоприятных мутаций. В итоге мощность лимитирующей структуры нарастает вдоль филетического маршрута как ряд:
 (5.4)
(5.4)
где Р 0— мощность структуры до начала действия движущего отбора; x i— число раз, в которое увеличивается мощность структуры в ходе фиксации i -й мутации. Это как раз те «незначительные изменения», аккумуляция которых, по выражению Дарвина, ведет к созданию органа «любой мыслимой степени совершенства».
Заметим, что логарифм мощности структуры (ln P ) после фиксации и благоприятных мутаций определяется равенством
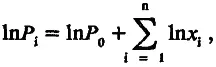 (5.5)
(5.5)
где стоящий под знаком суммы ln x i представляет собой вклад i -й благоприятной мутации в логарифм мощности рабочей структуры; для удобства дальнейшего изложения этот вклад будем обозначать а i'. Если единицей измерения мощности рабочей структуры выбрать ее исходную величину Р 0, то (учитывая, что lnl = 0) уравнение (5.5) можно представить в более простой форме:
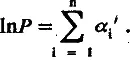 (5.6)
(5.6)
Напомним, что а i ' — вклад i -й мутации, зафиксированной отбором, направленным на усиление лимитирующей структуры.
Теперь введем одно определение: локусы, по которым шло замещение новых аллелей, повышающих мощность структуры в ее историческом развитии, назовем специальными генами структуры. Вполне очевидно, что некоторые из этих генов могут повторно мутировать, порождая новые аллели, усиливающие лимитирующую функцию. Таким образом, среди n благоприятных мутаций, зафиксированных движущим отбором, далеко не все связаны с увеличением числа специальных генов N.
Представим себе, что изменение среды приняло противоположное направление. В новых условиях мощность структуры, развитой за предыдущий период, может стать избыточной, поэтому ничто не будет препятствовать накоплению мутаций, ослабляющих экспрессию ее специальных генов. Более того, энергия и вещество, которые ранее расходовались на поддержание высокой мощности этой структуры, теперь могут быть направлены на создание дополнительной биомассы, т. е. на повышение абсолютной приспособленности вида. Таким образом, ослабление рабочей структуры в данном случае представляет собой адаптивный ответ вида на «требование» изменившейся среды. Через некоторое время новая среда может снова потребовать усиления функции той же структуры, и снова благоприятными станут мутации, усиливающие экспрессию ее специальных генов.
Таким образом, для вида, предки которого прошли через многократные изменения требований среды к интенсивности функции определенной рабочей структуры, логарифм ее мощности можно представить в виде суммы вкладов N специальных генов, где вклад каждого такого гена (его экспрессия ) ( а ) может изменяться от нуля до какого-то максимального значения, т. е.
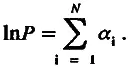 (5.7)
(5.7)
Это выражение удобнее представить в несколько иной форме:
ln P = Na, (5.8)
где а — уровень средней экспрессии специального гена, т. е. его средний вклад в логарифм мощности рабочей структуры. У разных видов а может варьировать в очень широком диапазоне: от нуля, когда все специальные гены «выключены», до величины, соответствующей уровню их максимальной активности. Ясно, что уровень средней экспрессии специального гена рабочей структуры зависит от предыстории вида, от того, как долго среда «требовала» от предков усиления мощности структуры и как долго была к ней безразлична или даже «ждала» ослабления.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Эволюция и прогресс»
Представляем Вашему вниманию похожие книги на «Эволюция и прогресс» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.









Обсуждение, отзывы о книге «Эволюция и прогресс» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.