Владимир Бердников - Эволюция и прогресс
Здесь есть возможность читать онлайн «Владимир Бердников - Эволюция и прогресс» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Новосибирск, Год выпуска: 1991, ISBN: 1991, Издательство: Наука, Жанр: Биология, sci_popular, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Эволюция и прогресс
- Автор:
- Издательство:Наука
- Жанр:
- Год:1991
- Город:Новосибирск
- ISBN:5-02-030062-4
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Эволюция и прогресс: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Эволюция и прогресс»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Книга предназначена для биологов широкого профиля, а также всех интересующихся вопросами эволюции живых существ.
Эволюция и прогресс — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Эволюция и прогресс», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Пусть разница средних значений признака двух изогенных линий обусловлена отличием в силе аллелей только одного локуса. Тогда генотип линии Р 1 обозначим аа, а генотип линии Р 2— АА. Примем еще одно упрощающее условие: пусть по степени доминирования оба аллеля (а и А) равны (h = 0,5). Теперь введем представление об эффекте аллельного замещения. Будем считать, что замещение одного слабого аллеля на сильный увеличивает генотипическое значение признака на α единиц. Тогда замещение обоих слабых аллелей на сильные увеличит это значение на 2α единиц. Вспомним, что генотипическое значение признака равно его среднему значению у особей с одинаковым генотипом, т. е.
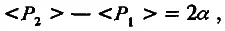 (4.6)
(4.6)
символ < > означает среднее значение признака в линии.
Проведя массовые скрещивания особей таких линий, получим популяцию гибридов F 1 :
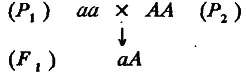
Все особи популяции F 1 имеют один и тот же генотип аА, поэтому изменчивость признака в данном случае обусловлена исключительно средой. Будем считать, что средовая дисперсия для всех популяций (F 1, P 1 и Р 2 ) одинакова и равна σ e 2. Среднее значение признака у особей F 1 (обозначим его т ) должно равняться генотипическому значению гетерозигот аА, т. е. оно должно на α единиц превосходить среднее значение особей линии P 1 и настолько же уступать среднему значению особей Р 2. Иными словами, m попадает точно в середину интервала между средними значениями признака обеих родительских популяций. Тогда средние значения признака всех рассмотренных популяций можно представить как
1> = m ; < Р 1 > = m — а; <���Р 2> = m + а . (4.7)
Итак, m, m — а и m + а — генотипические значения признака у особей с генетической конституцией аА, аа и АА соответственно.
Перейдем к популяции F 2 , возникшей или при самооплодотворении, или при панмиктическом скрещивании особей из F 1 :
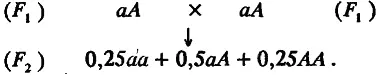
Из этой схемы видно, что вся совокупность особей F 2 разбивается на три генотипических класса; каждый из них можно охарактеризовать его долей в выборке и средним значением признака. Одна четверть особей имеет генотип аа, другая четверть — генотип АА и половина — генотип аА. Поскольку средние значения признака у особей с этими генотипами равны соответственно m — а, m + а и m, то по (4.3) и (4.4) легко рассчитать среднее значение (М) и дисперсию (σ g 2) для распределения особей по генотипическим классам:
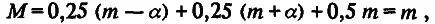 (4.8)
(4.8)
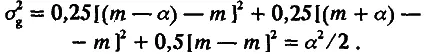 (4.9)
(4.9)
Таким образом, популяция F 2 обладает дисперсией ( а 2 /2), обусловленной различием особей по генотипу. Кроме того, из-за «шума» среды популяция обладает и средовой дисперсией σ e 2. Этот шум не сдвигает средних значений, поэтому 2> = m. Эффекты среды и генотипа независимы, отсюда следует, что дисперсия по признаку в поколении F 2 должна быть больше средовой на положительную величину а 2 /2, т. е.
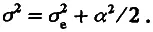 (4.10)
(4.10)
Теперь попробуем рассмотреть более общий случай, когда особи двух изогенных линий различаются аллелями n несцепленных локусов. По-прежнему будем считать, что все слабые аллели собраны у линии P 1, а все сильные — у линии Р 2. Проведем их скрещивание:
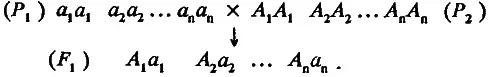
При оценке среднего значения популяции F 1 сделаем два предположения: во-первых, по степени доминирования все аллели равны и, во-вторых, замещение в каждом локусе одного слабого аллеля на сильный увеличивает генотипическое значение признака на одну и ту же величину а. Следовательно, разность средних значений родительских популяций должна быть равна 2 na, а среднее значение признака в популяции F, (обозначим его 1> ) будет находиться в точке m, т. е. точно посередине между средними значениями родительских линий. Такая модель, где вклады всех аллелей в величину признака суммируются, получила название аддитивной. Главным основанием для ее применения является попадание среднего значения признака в популяциях F 1 и F 2 посередине между средними значениями родительских популяций. Итак, для случая n локусов
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Эволюция и прогресс»
Представляем Вашему вниманию похожие книги на «Эволюция и прогресс» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.









Обсуждение, отзывы о книге «Эволюция и прогресс» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.