Денис Соломатин - Математические модели в естественнонаучном образовании. Том II
Здесь есть возможность читать онлайн «Денис Соломатин - Математические модели в естественнонаучном образовании. Том II» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Год выпуска: 2022, Жанр: Детская образовательная литература, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Математические модели в естественнонаучном образовании. Том II
- Автор:
- Жанр:
- Год:2022
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Математические модели в естественнонаучном образовании. Том II: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Математические модели в естественнонаучном образовании. Том II»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Математические модели в естественнонаучном образовании. Том II — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Математические модели в естественнонаучном образовании. Том II», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
В работе Канна 1987 года была впервые предпринята попытка определить местонахождение митохондриальной Евы в Африке. Поддерживая теорию происхождения человека «из Африки», было построено дерево с корнями, которое, как утверждается, показывает отношения между 147 людьми. Сколько топологически различных деревьев нужно было бы рассмотреть, если бы действительно рассматривалась каждая возможность? Возможно, для ответа на этот вопрос придется использовать формулу Стирлинга: 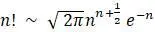 . Здесь символ «∼» можно интерпретировать как «приблизительно». Изучению последствий трудности рассмотрения стольких деревьев посвятил свою работу Гиббонс в 1992 году.
. Здесь символ «∼» можно интерпретировать как «приблизительно». Изучению последствий трудности рассмотрения стольких деревьев посвятил свою работу Гиббонс в 1992 году.
5.1.8. Филогенез четырех терминальных таксонов A, B, C и D связан по определенному метрическому дереву. Суммарные расстояния между таксонами вдоль дерева оказались такими же, как в таблице 5.1.
Таблица 5.1. Расстояния между таксонами для задачи 5.1.8
A B C D
A .6 .6 .2
B .4 .6
C .6
а. Используя любой подход, который пожелаете, определите правильное некорневое дерево, относящееся к этим таксонам, а также все длины его ребер. Объясните, как исключили другие топологические деревья.
б. Можете ли определить корень дерева по этим данным? Объясните, почему да или почему нет.
Примечание: Методы решения такого рода проблем являются предметом следующих разделов.
5.2. Построение дерева дистанционными методами UPGMA и FM
При построении филогенетического дерева таксоны, которые хотим связать, обычно являются теми, которые живут в настоящее время. Есть информация, такая как последовательности ДНК, от терминальных таксонов и нет информации от тех, которые представлены внутренними вершинами. В действительности, даже не знаем, какие внутренние вершины должны существовать, потому что не знаем даже топологию дерева.
Первым классом методов построения филогенетических деревьев, которые обсудим, являются дистанционные методы. Они пытаются построить дерево, используя информацию, которая предположительно описывает общие расстояния между терминальными таксонами вдоль дерева.
Чтобы понять, как получить эти расстояния, представьте, что пытаемся найти эволюционные отношения четырех видов: 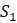 ,
, 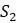 ,
, 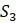 и
и 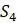 . Выбирая тот или иной ортологичный участок ДНК из их геномов, получаем и выравниваем последовательности из каждого. Если модель замены оснований Джукса-Кантора, рассмотренная в главе 4, кажется подходящей для имеющихся данных, то вычисляем расстояния Джукса-Кантора между каждой парой последовательностей. Получатся оценки расстояний по дереву, которые сводим в Таблицу 5.2.
. Выбирая тот или иной ортологичный участок ДНК из их геномов, получаем и выравниваем последовательности из каждого. Если модель замены оснований Джукса-Кантора, рассмотренная в главе 4, кажется подходящей для имеющихся данных, то вычисляем расстояния Джукса-Кантора между каждой парой последовательностей. Получатся оценки расстояний по дереву, которые сводим в Таблицу 5.2.
В зависимости от данных последовательности могли бы вместо этого принять другую модель подстановки оснований, что привело бы к использованию другой формулы расстояния, такой как в 2-параметрической модели Кимуры или логарифмическое расстояние. Несмотря на это, расстояние, которое вычисляем между последовательностями, считается мерой количества произошедших мутаций. Если бы эти расстояния были точной мерой количества произведенных мутаций, они бы соответствовали между конечными таксонами в найденном метрическом дереве.
Таблица 5.2. Расстояния между таксонами
.45 .27 .53
.40 .50
.62
На самом деле даже не ожидаем найти дерево, которое точно соответствует имеющимся данным; в конце концов, расстояния выводятся из данных последовательности и не должны быть точно правильными. Более того, метод вывода расстояний зависел от модели, которая включала дополнительные предположения, которые, безусловно, не встречаются в реальных организмах. Надеемся, однако, что построенное дерево не будет слишком чувствительно к такого рода ошибкам на больших расстояниях.
Первый метод, который рассматриваем, называется методом среднего расстояния или, более формально, невзвешенным парно-групповым методом с арифметическими средними (UPGMA). Этот метод создает корневое дерево и предполагает наличие молекулярных часов. Самый простой способ понять алгоритм – это ознакомиться с примером его использования.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Математические модели в естественнонаучном образовании. Том II»
Представляем Вашему вниманию похожие книги на «Математические модели в естественнонаучном образовании. Том II» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Математические модели в естественнонаучном образовании. Том II» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.