Роберт Немов - Психология. Книга 1
Здесь есть возможность читать онлайн «Роберт Немов - Психология. Книга 1» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: Старинная литература, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.
- Название:Психология. Книга 1
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:4.5 / 5. Голосов: 2
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Психология. Книга 1: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Психология. Книга 1»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Психология. Книга 1 — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Психология. Книга 1», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
состояние возбуждения при стимулировании всего поля сетчаточных рецепторов определенной
конфигурацией, например линией или краем специфической ориентации и наклона. Они не
возбуждаются, если стимулируется не весь «клеточный ансамбль», а отдельные палочки и
колбочки отдельными световыми точками в той же области сетчатки. Эти клетки, чувствительные
к краям и линиям, расположены в сенсорной проекционной зоне зрения.
В заключение обсуждения нервных механизмов зрения отметим, что вклад в их работу вносит уже
отмеченная нами функциональная асимметрия мозга. Оба полушария, левое и правое, играют
различную роль в восприятии и формировании образа. Для правого полушария характерны
высокая скорость работы по опознанию, его точность и четкость. Такой способ опознания
предметов можно определить как интегрально-синтетический, целостный по преимуществу,
структурно-смысловой2. Правое полушарие, вероятно, производит сличение образа с некоторым
имеющимся в памяти эталоном на основе вьщеления в воспринимаемом объекте некоторых
информативных признаков. С помощью же левого полушария осуществляется в основ-
1Хохберг Дж. Единицы мозговой активности, клеточные ансамбли и рецептивные поля //
Хрестоматия по ощущению и восприятию. — М., 1975. — С. <���п.
2См.: Николаекко КН., Черниговская Т.В. Опознание сложных цветовых образов и функциональная
асимметрия мозга // Вопросы психологии. — 1989. — № 1.
60
ном аналитический подход к формированию образа, связанный с последовательным перебором
его элементов по определенной программе. Но левое полушарие, работая изолированно, по-
видимому, не в состоянии интегрировать воспринятые и выделенные элементы в целостный образ.
С его помощью производится классификация явлений и отнесение их к определенной категории
через обозначение словом. Таким образом, в восприятии с разными функциями одновременно
принимают участие оба полушария головного мозга.
Рассмотрим теперь физиологические механизмы
внимания.
Как общее состояние
сосредоточенности оно связано с повышением возбудимости к.г.м. в целом или ее отдельных
участков. Это в свою очередь соотносится с активностью отдельных частей ретикулярной
формации. Те ее отделы, которые своей деятельностью порождают общий эффект возбуждения,
входят в структуры, связанные с ориентировочным рефлексом, автоматически возникающим при
любых неожиданных и заметных изменениях стимулов, воздействующих на организм. В свою
очередь те отделы ретикулярной формации, которые вызывают специфический эффект
возбуждения, функционируют, по-видимому, в рамках анатомо-физиологической системы
доминанты. С ней же скорее всего соотносим в своем действии и избирательный механизм
регуляции внимания через актуализацию потребностей, а также механизм волевого управления
вниманием через кортикально-подкорковые связи.
Много споров в науке было вокруг вопроса о физиологических основах внимания. Органическая
интерпретация процессов, внимания привлекала исследователей на протяжении всего времени его
изучения. Т.Рибо одним из первых пытался представить физиологическую схему произвольного
внимания, связанного с волевым усилием, возникающим в процессе припоминания (рис. 22).
Рассмотрим его концепцию.
На рис. 22 М — центр, где хранятся воспоминания о некогда совершенных действиях
(предполагаемый центр долговременной памяти на движения); Si и S2 — центры, где сохраняются
сенсорные образы (центры сенсорной памяти). Те и другие локализованы в коре головного мозга,
представленной на рисунке дугообразной заштрихованной областью. Эти центры между собой
соединены волокнами ассоциативного типа; R — центр, расположенный ниже коры и связанный с
регуляцией движений; С,, С2, С3 — совокупность сенсорных центров разного уровня, передающих
коре больших полушарий информацию о со-
61
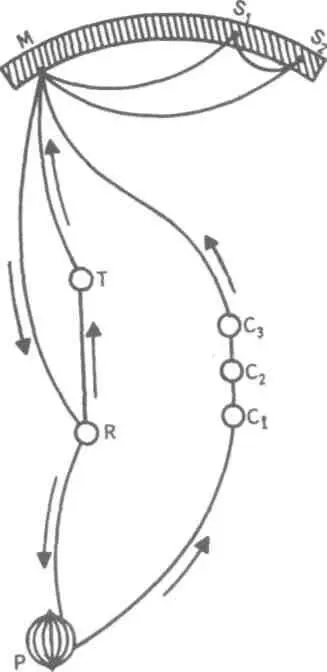
стоянии мышечной системы М («мышечное чувство», по Т.Ри-бо); Т — область таламуса.
Допустим, что в кортикальных центрах М, Sx и S2 существует некоторый уровень возбуждения, т.е.
они находятся в состоянии, связанном с вниманием, субъективно представляя собой ряд
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Психология. Книга 1»
Представляем Вашему вниманию похожие книги на «Психология. Книга 1» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.










Обсуждение, отзывы о книге «Психология. Книга 1» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.