Charles J. Dorman - Structure and Function of the Bacterial Genome
Здесь есть возможность читать онлайн «Charles J. Dorman - Structure and Function of the Bacterial Genome» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.
- Название:Structure and Function of the Bacterial Genome
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Structure and Function of the Bacterial Genome: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Structure and Function of the Bacterial Genome»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Structure and Function of the Bacterial Genome Blends knowledge of gene regulatory mechanisms with a consideration of nucleoid structure and dynamics Offers a 'DNA-centric' approach to considering transcription control Views horizontal gene transfer from a gene regulation perspective Assesses the opportunities and limitations of designing synthetic microbes or rewiring existing ones
is an ideal book for graduate and undergraduate students studying microbial cell biology, bacterial pathogenesis, gene regulation, and molecular microbiology. It will also appeal to principal investigators conducting research on these and related topics and researchers in synthetic biology and other arms of biotechnology.
Structure and Function of the Bacterial Genome — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Structure and Function of the Bacterial Genome», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
1.33 Transcription and Nucleoid Structure
Several investigations have made links between patterns of transcription and the superstructure of the bacterial nucleoid. At a practical level, replication fork movement must be reconciled with the needs of transcription (initiation, elongation, and termination), so aligning replisome movement with the direction of gene transcription avoids significant conflicts between DNA and RNA polymerases. Collisions between the replisome and RNA polymerase are known to cause severe inhibition of replisome progression (Mirkin and Mirkin 2005). Transcriptional promoters that oppose the direction of replisome movement serve to pause the replication fork, while transcription terminators that are aligned with the direction of replisome movement also act as replication fork pause sites (Mirkin et al. 2006). Conflicts between the replisome and RNA polymerase can generate R‐loops, stalling replication, and transcription in the affected region until RNase H removes the R‐loop (Kuziminov 2018). Unresolved R‐loops also result in hyper‐recombination and genome instability, so avoiding replication–transcription conflicts is very desirable ( Figure 1.16).
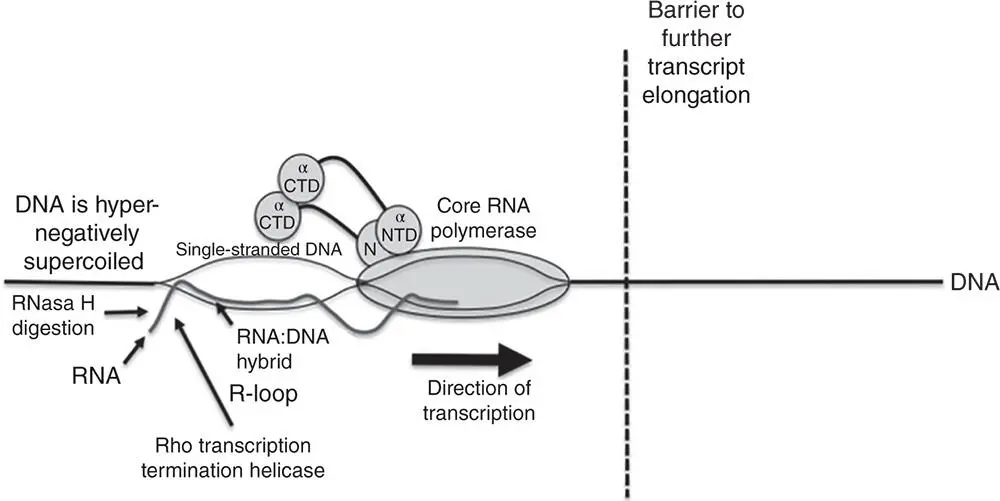
Figure 1.16R‐loop formation. When RNA polymerase reads a G+C‐rich template, stalls and backtracks, it leaves a domain of hypernegatively supercoiled in its wake and the associated transcription stalling may allow the RNA transcript to base pair with its DNA template strand, leaving the non‐transcribed strand as a single‐stranded bubble. Other impediments to RNA polymerase progression include head‐on collisions with other transcription units or with replisomes (the barrier is represented by the vertical gapped line). Loss of topoisomerase I activity is known to promote R‐loop formation because it encourages the accumulation of hyper negative superhelicity in DNA that is being transcribed (or replicated). Failure to process and remove RNA loops can lead to DNA damage, including double‐stranded breaks and hyper‐recombination. The Rho transcription‐terminating helicase (Figure 3.4) helps to suppress R‐loop formation by preventing backtracking by RNA polymerase while RNase H eliminates R‐loops by removing the RNA component of the RNA:DNA hybrid in R‐loops.
A correlation has also been reported between gene essentiality and alignment with the direction of replication fork movement (Rocha and Danchin 2003). Overall, one finds more genes on the leading strand than on the lagging strand of the chromosome and this may reflect the outcome of evolutionary pressure to minimise collisions (Rocha 2008). The question of replication and transcription alignment/collision also has a DNA topological dimension, in that converging polymerases will create, and trap between them, a domain of positively supercoiled DNA that must be resolved by type II topoisomerases: gyrase or topo IV (Crisona et al. 2000; Kato et al. 1992). The mechanisms of action of these enzymes bring an increased risk of double‐stranded breaks occurring in the chromosome with potentially lethal consequences for the cell (Hiasa et al. 1996; Lockshon and Morris 1983). These factors may impose limits on the options for gene orientation on the chromosome, influencing its evolution.
Experiments using chromosome conformation capture (3C) methods (Dekker et al. 2002) have suggested a link between DNA replication, transcription, and the organisation of the bacterial nucleoid (Cagliero et al. 2013; Le et al. 2013; Le and Laub 2016; Lioy et al. 2018; Meyer et al. 2018). Work that exploited site‐specific recombination as a measure of interaction frequency within the bacterial genome suggested that transcription activity and the associated changes in local DNA topology have a structural function in the nucleoid (Booker et al. 2010; Higgins 2014). Furthermore, the global level of DNA supercoiling in the nucleoid can be tuned by treating the bacterium with rifampicin, an inhibitor of RNA polymerase (Rovinskiy et al. 2012). Experiments with trimethylpsoralen crosslinking have provided evidence that a gradient of DNA supercoiling exists extending from the origin of chromosome replication to the Ter macrodomain (Lal et al. 2016). Bioinformatic and experimental studies show that the distribution of binding sites around the chromosome for DNA gyrase is non‐uniform, with more sites being detected close to the origin of replication (Jeong et al. 2004; Sobetzko et al. 2012; Sutormin et al. 2019). This could indicate that the Ori‐proximal part of the chromosome is the most underwound, contradicting the data from the psoralen‐binding‐and‐crosslinking studies (Lal et al. 2016). On the other hand, it may indicate a need to compensate for the paucity of negative supercoiling in the Ori‐proximal part of the chromosome that is predicted by those studies. Further evidence that the chromosome is not uniformly supercoiled has come from investigations in which supercoiling‐sensitive genes have been placed at different locations in the genome (Bryant et al. 2014). A comprehensive survey of the effects of gene position on transcription in E. coli showed that the propensity for transcription varies with chromosomal location: horizontally acquired genetic elements are associated with quiescent regions while ribosomal and other metabolic genes are in highly active zones (Scholz et al. 2019). It should be noted, however, that several studies which have explored the influence of gene position (including the possible contribution of differences in local DNA supercoiling) did not detect clear changes in the level of expression of the test gene(s) at different sites around the chromosome (Block et al. 2012; Brambilla and Sclavi 2015; Chandler and Pritchard 1975; Miller and Simons 1993; Pavitt and Higgins 1993; Schmid and Roth 1987; Sousa et al. 1997; Thompson and Gasson 2001; Ying et al. 2014).
1.34 Nucleoid‐associated Proteins (NAPs) and Nucleoid Structure
Sequence‐dependent DNA‐binding proteins such as MaoP, MatP, SeqA, and SlmA play important roles in the organisation of the chromosome in the nucleoid. Yet the term ‘nucleoid‐associated protein’, or NAP, is usually reserved for another group of DNA‐binding proteins, not all of which are sequence‐dependent for DNA binding. Most NAPs were discovered in roles other than organising the nucleoid. Several were first encountered as contributors to the efficient operation of site‐specific recombination systems in bacteria or their phage (reviewed in Dorman and Bogue 2016). Later, their more general contributions to cell physiology became appreciated. We have already encountered two, FIS and IHF, as components of the systems that govern the initiation of chromosome replication ( Section 1.3). Originally, FIS was identified as an enhancer‐binding protein that improved the efficiency of the DNA inversion events responsible for phase‐variable expression of tail fibre genes in bacteriophage Mu, and for flagellar phase variation in Salmonella (Koch and Kahmann 1986; Johnson et al. 1986). IHF was detected as an essential factor for the site‐specific entry of the bacteriophage lambda genome into the att λ site on the chromosome of E. coli (Nash and Robertson 1981) ( Section 1.35). These and other NAPs have emerged as important regulators of a multitude of bacterial genes (Dillon and Dorman 2010). They have mechanisms of action that typically involve the making of adjustments to local DNA architecture. In many cases, these adjustments affect gene expression at the level of transcription (or beyond) and include direct or indirect effects on nucleoid structure. A summary of the key features of some of the most important (i.e. best‐studied) NAPs is given in the following sections.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Structure and Function of the Bacterial Genome»
Представляем Вашему вниманию похожие книги на «Structure and Function of the Bacterial Genome» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Structure and Function of the Bacterial Genome» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.