Charles J. Dorman - Structure and Function of the Bacterial Genome
Здесь есть возможность читать онлайн «Charles J. Dorman - Structure and Function of the Bacterial Genome» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.
- Название:Structure and Function of the Bacterial Genome
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Structure and Function of the Bacterial Genome: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Structure and Function of the Bacterial Genome»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Structure and Function of the Bacterial Genome Blends knowledge of gene regulatory mechanisms with a consideration of nucleoid structure and dynamics Offers a 'DNA-centric' approach to considering transcription control Views horizontal gene transfer from a gene regulation perspective Assesses the opportunities and limitations of designing synthetic microbes or rewiring existing ones
is an ideal book for graduate and undergraduate students studying microbial cell biology, bacterial pathogenesis, gene regulation, and molecular microbiology. It will also appeal to principal investigators conducting research on these and related topics and researchers in synthetic biology and other arms of biotechnology.
Structure and Function of the Bacterial Genome — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Structure and Function of the Bacterial Genome», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Macrodomains play important roles in determining the choreography of the daughter chromosomes, as these segregate prior to cell division (Espéli et al. 2008). They also correlate with global gene expression patterns, suggesting that the overall gene expression programme of the cell is written into the architecture of the nucleoid (Cameron et al. 2017; Sobetzko 2016; Sobetzko et al. 2012). To appreciate the significance of the connections between nucleoid structure and gene expression, it will be necessary to consider the contributions made to both by variable DNA structure and NAPs.
1.26 DNA in the Bacterial Nucleoid
DNA in bacterial cells is maintained in an underwound state and this affects the shape that the DNA duplex adopts as it seeks to adopt a minimal energy conformation. The underwinding arises due to a deficit in helical turns, i.e. the number of times the two DNA strands twist around the duplex axis. The twist deficiency imparts torsional stress to the duplex, which is relieved by allowing the duplex to adopt a writhed confirmation in which the helical axis coils around itself. This coiling of the already coiled DNA duplex creates supercoiling and has the effect of making the DNA molecule more compact. In the context of the nucleoid, such compaction assists with solving the problem of packaging the genetic material within the cell. The most supercoiled parts of the chromosome form branches, facilitating further compaction.
1.27 DNA Topology
The topological state of a DNA molecule is described by three parameters: the linking number (the number of times one DNA strand crosses the other in the duplex); the twist (the number of complete turns made by the strands around one another along the length of the duplex); and the writhe (the number of writhing turns made by the duplex axis around itself) (Bauer et al. 1980; Boles et al. 1990; Vinograd et al. 1965). Changes to the linking number can be made by breaking one or both strands of the duplex, twisting the DNA with or against the sense of the double helix, and then resealing the strand breaks (Higgins and Vologodskii 2015). Twisting the DNA in keeping with the sense of the double helix tightens the duplex and imparts positive writhe, resulting in positive supercoiling. Twisting the DNA against the sense of the double helix underwinds the molecule, imposing negative writhe and therefore negative supercoiling (Sinden 1994). In nature, negative supercoiling is the norm although positive supercoils do arise naturally, especially during the movement of the DNA replication fork ( Figure 1.14) and during transcription (Liu and Wang 1987) ( Figure 1.15). In contrast to mesophilic bacteria like E. coli , hyperthermophilic archaea that live in environments that are characterised by very high temperatures possess a reverse gyrase activity and maintain their DNA in a positively supercoiled state (Couturier et al. 2014; Lipscomb et al. 2017). Reverse gyrase, first described in Sulfolobus acidocaldarius in 1984, uses ATP to introduce positive supercoils into DNA (Kikuchi and Asai 1984; Ogawa et al. 2015).
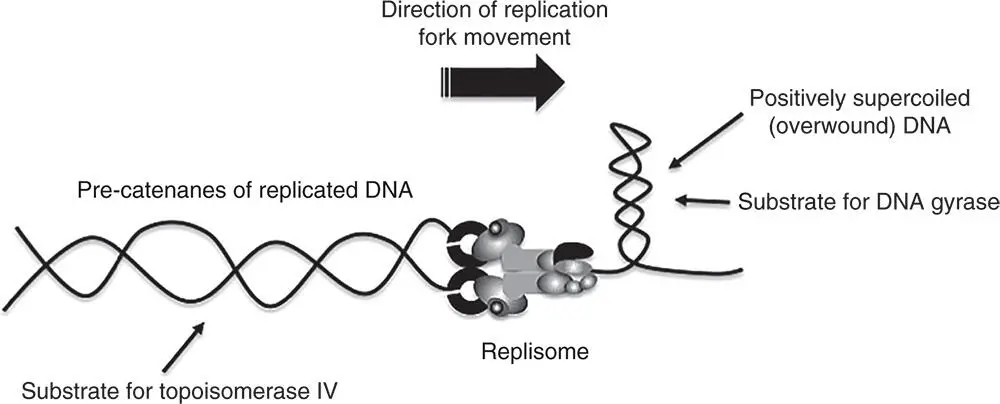
Figure 1.14The topological consequences of replisome activity. The replication fork, moving from left to right, over winds the DNA ahead to create a domain of positive supercoiling. Unless this is relaxed, the replisome will be unable to advance further. Relaxation of the positively supercoiled domain is carried out by DNA gyrase: it neutralises the positive supercoils by introducing negative ones. In C. crescentus , the GapR protein binds to positively supercoiled DNA and stimulates its relaxation by type II topoisomerases (see Guo et al. 2018). The newly synthesised DNA behind the replisome becomes catenated. These interwound, double‐stranded DNA molecules are decatenated by DNA topoisomerase IV, allowing the daughter chromosomes to be segregated at cell division.
For further information, see Lopez et al. (2012) and Postow et al. (2001).
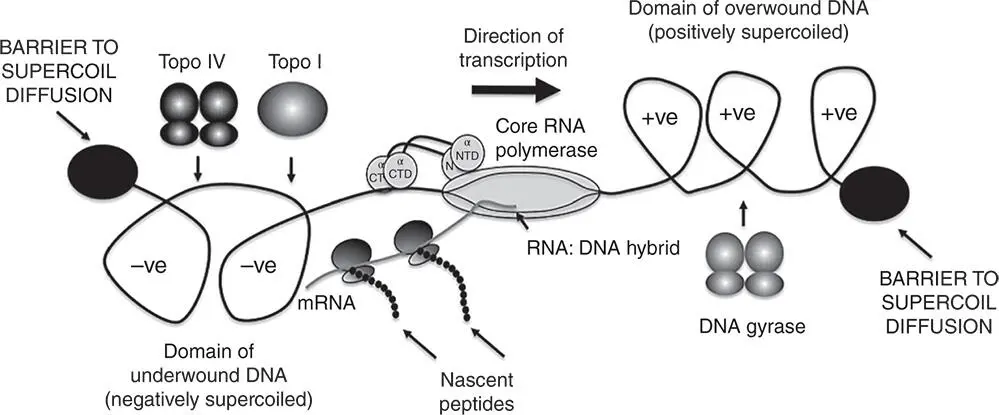
Figure 1.15The topological consequences of transcription. This is the twin supercoil domain model, as proposed by Liu and Wang (1987) and supported by numerous independent experiments. Core RNA polymerase is engaged in transcript elongation and the mRNA is simultaneously being translated by polyribosomes to produce nascent peptides. As the coupled transcription‐translation complex moves from left to right, the DNA template ahead becomes overwound, or positively supercoiled, while the DNA behind becomes underwound, or negatively supercoiled. This situation will bring transcription to a halt as the machinery jams. This is because: (a) the domains of supercoiled DNA cannot be resolved by supercoil lateral diffusion because the transcribed region is bounded by topological barriers (solid black discs) and (b) the bulky transcription‐translation complex cannot rotate around the DNA to relieve the torsional tension in the DNA duplex. The solution to the problem is provided by the DNA‐relaxing activities of topoisomerases: DNA gyrase removes the positive supercoils by introducing negative supercoils, while the negatively supercoiled domain is relaxed by DNA topoisomerase I and IV. Interference with these relaxation processes can result in undesirable outcomes, such as R‐loop formation ( Figure 1.16). Topological barriers can be created by head‐to‐head transcription collisions and by collisions between converging replisomes and RNA polymerases; they can also arise from the presence of nucleoprotein complexes and distortions (e.g. sharp bends) in the DNA duplex.
A covalently closed, circular duplex DNA molecule that is neither overwound nor underwound is said to be topologically relaxed. If this circular duplex undergoes an increase in its linking number (overwinding) or a decrease (underwinding) it retains an identical nucleotide sequence compared with the relaxed form but differs from this form topologically (Sinden 1994). For this reason, the relaxed, overwound, and underwound isomeric forms of the circular duplex are referred to as topoisomers of the same DNA molecule. Enzymes that produce topological changes in DNA by altering the linking number are called topoisomerases and E. coli has four: topoisomerase I (topo I), DNA gyrase (a topo II family member), topoisomerase III (topo III), and topoisomerase IV (topo IV) (Bates and Maxwell 2005; Wang 2002) ( Table 1.1).
Table 1.1The topoisomerases of E. coli .
| Enzyme name (typea) | Molecular mass (kDa) | Gene(s) | Comments |
| (Type I) | |||
| Topoisomerase I | 97 | topA | DNA swivelase that makes a transient cut in one strand of the DNA duplex, binds to the cut site via a 5′‐phosphotyrosine bond; relaxes negatively supercoiled DNA; requires Mg 2+ |
| Topoisomerase III | 73.2 | topB | Relaxes negatively supercoiled DNA; decatenase; has catenase activity in association with RecQ; requires Mg 2+ |
| (Type II) | |||
| DNA gyrase (Topoisomerase II) | 105 (A subunit) 95 (B subunit) | gyrA (A subunit) gyrB (B subunit) | ATP‐dependent negative supercoiling activity; relaxes negative supercoils in an ATP‐independent manner; relaxes positively supercoiled DNA; binds DNA transiently via a 5′‐phosphotyrosine bind; requires Mg 2+ |
| Topoisomerase IV | 75 (ParC) 70 (ParE) | parC (ParC; GyrA‐like) parE (ParE; GyrB‐like) | Decatenase activity; interacts with MukBEF; relaxes negative supercoils; requires ATP, Mg 2+ |
aType I enzymes change the linking number of the duplex DNA substrate in steps of 1 (ΔLk = 1) while type II enzymes change the linking number in steps of 2 (ΔLk = 2).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Structure and Function of the Bacterial Genome»
Представляем Вашему вниманию похожие книги на «Structure and Function of the Bacterial Genome» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Structure and Function of the Bacterial Genome» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.