Jane Flint - Principles of Virology
Здесь есть возможность читать онлайн «Jane Flint - Principles of Virology» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.
- Название:Principles of Virology
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Principles of Virology: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Principles of Virology»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Volume I: Molecular Biology
Volume II: Pathogenesis and Control
Principles of Virology, Fifth Edition
Principles of Virology — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Principles of Virology», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
The nuclear pore complex allows passage of cargo in and out of the nucleus by either passive diffusion or facilitated translocation. Passive diffusion does not require interaction between the cargo and components of the nuclear pore complex and becomes inefficient as molecules approach 9 nm in diameter. Objects as large as 39 nm in diameter can pass through nuclear pore complexes by facilitated translocation via specific interactions between the cargo and components of the nuclear pore complex. Many subviral particles are too large to pass through the nuclear pore complex, but several strategies overcome this limitation.
Nuclear Localization Signals
Proteins that reside within the nucleus are characterized by the presence of specific nuclear targeting sequences. Such nuclear localization signalsare both necessary for nuclear entry of the proteins in which they are present and sufficient to direct heterologous, nonnuclear proteins to enter this organelle. Nuclear localization signals identified by these criteria share a number of common properties: they are generally fewer than 20 amino acids in length, and are usually rich in basic amino acids. Although no consensus nuclear localization sequence can be defined, most nuclear localization signals belong to one of two classes, simple or bipartite sequences ( Fig. 5.25). A particularly well-characterized example of a simple nuclear localization signal is that of simian virus 40 large T antigen, which comprises five contiguous basic residues flanked by hydrophobic amino acids.

Figure 5.24 Structure and organization of the nuclear pore complex. (Bottom left) Cartoon depiction of the nuclear membrane, showing the topology of the nuclear pore complexes. The entire structure of the nuclear pore complex of Saccharomyces cerevisiae , comprising 552 proteins, was determined using cryo-electron tomography and modeling (see Kim SJ et al. 2018. Nature 555:475–482). The yeast and human nuclear pore complexes are highly conserved. (Top) Side view of the nuclear pore showing three spokes. Spokes are high-ordered subcomplexes formed by NUPs (nucleoporins). Spokes in turn assemble into larger complexes to form two outer and two inner rings flanking a membrane ring. The complex is completed by a nuclear basket composed of NUPs. The outer ring comprises export complexes (olive green) and connectors (tan) with the inner ring (magenta, pink, and green). The inner rings sandwich the membrane ring (light yellow). The nucleoplasmic outer ring links to the nuclear basket (only partly depicted as light blue filaments). (Bottom right) Top view of the nuclear pore from the cytoplasm showing a model of the central transporter (green). Structure reconstructions courtesy of Michael Rout, Rockefeller University, New York.
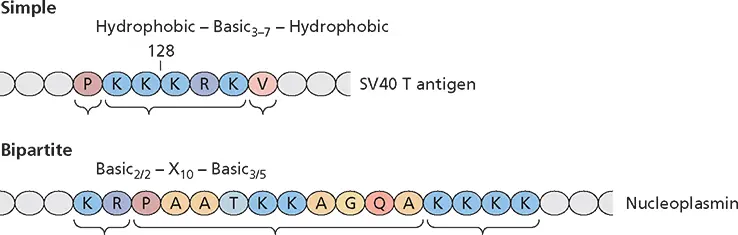
Figure 5.25 Nuclear localization signals. The general form and specific examples of simple and bipartite nuclear localization signals are shown in the one-letter amino acid code, where X is any residue. Bipartite nuclear targeting signals are defined by the presence of two clusters of positively charged amino acids separated by a spacer region of variable length and sequence. Both clusters of basic residues, which often resemble the simple targeting sequences of proteins like simian virus 40 T antigen, are required for efficient import of the proteins in which they are found. The subscript indicates either length (3–7) or composition (e.g., 3/5 means at least 3 residues out of 5 are basic). Gold particles with diameters as large as 26 nm are readily imported following their microinjection into the cytoplasm, as long as they are coated with proteins or peptides containing a nuclear localization signal.
Import of a protein into the nucleus via nuclear localization signals occurs in two distinct, and experimentally separable, steps ( Box 5.5). A protein containing such a signal first binds to a soluble cytoplasmic receptor protein. This complex then engages with the cytoplasmic surface of the nuclear pore complex, a reaction often called docking, and is then translocated through the nuclear pore. In the nucleus, the complex is disassembled, releasing the protein cargo.
Nuclear Import of RNA Genomes
Influenza virus is among the few RNA-containing viruses with genomes that are replicated in the cell nucleus. The influenza virus genome, which consists of eight segments, is uncoated in the cytoplasm. After vRNPs separate from M1 and are released into the cytosol, they are imported rapidly into the nucleus ( Fig. 5.26A). Import depends on the presence of a nuclear localization signal in the NP protein, a component of vRNP: naked viral RNA does not dock onto the nuclear pore complex, nor does it enter the nucleus.
BOX 5.5
BACKGROUND
Transport through the nuclear pore
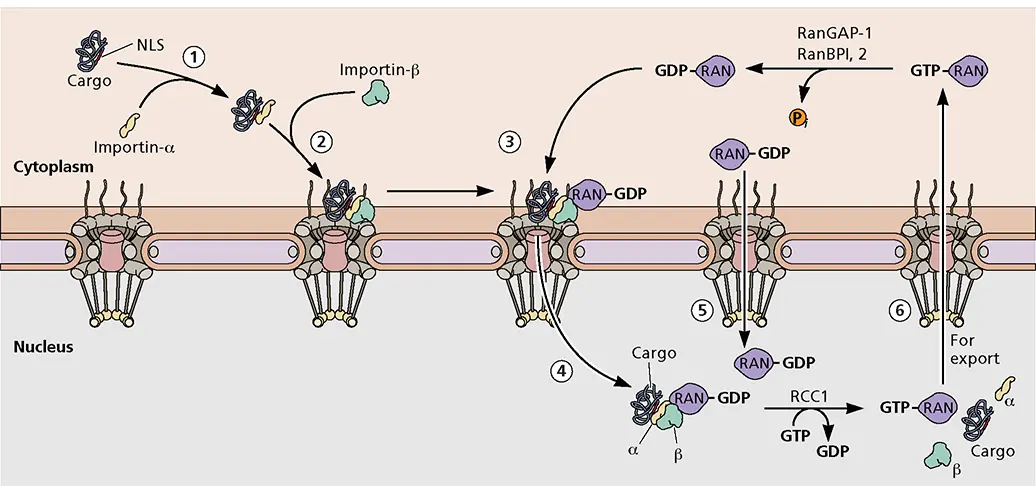
Schematic illustration of a classical nuclear import pathway.Data from Yang Q et al. 1998. Mol Cell 1:223–234.
Different groups of proteins are imported by specific nuclear transport receptor complexes. In what is known as the “classical system” of import, cargo proteins containing basic nuclear localization signals (NLS) bind to the cytoplasmic nuclear localization signal receptor protein importin-α (step 1 in the figure). This complex then binds importin-β1, which mediates docking with the nuclear pore complex by binding to members of a family of nucleoporins (step 2). It is likely that initial association involves nucleoporins present in the cytoplasmic filaments of the nuclear pore. Importin-β1 also interacts with RAN, a small RAS-related nucleotide-binding protein. RAN GTPase (step 3) is required for translocation of the complex into the nucleus through the central channel of the nuclear pore (step 4).
A single translocation through the nuclear pore complex does not require energy consumption. However, maintenance of a gradient of the guanosine nucleotide-bound forms of RAN is absolutely essential for continued transport. A RAN-specific guanine nucleotide exchange protein named RCC1 (regulator of chromosome condensation 1) resides in the nucleus and promotes the exchange of GDP to GTP. In contrast, a RAN-GTPase-activating protein (RANGAP-1) localized in the cytoplasm promotes hydrolysis of GTP. The nuclear pool of RAN-GDP is replenished by the action of nuclear transport factor 2 (NTF2), which transports RAN-GDP from the cytoplasm to the nucleus efficiently (step 5), where it can be converted to RAN-GTP. The asymmetric distribution of RCC1 and RANGAP-1 allows for the formation of a gradient of RAN-GTP/RAN-GDP. This gradient provides the driving force and directionality for nuclear transport.
Importin-β1 has a higher affinity for RAN-GTP, which is more abundant in the nucleus, than for RAN-GDP. Therefore, following import into the nucleus, importin-β1 binds to RAN-GTP and the complex disassembles, eventually releasing the cargo protein. The importin-β1 recycles to the cytoplasm bound to RAN-GTP (step 6). There, it is displaced by the action of two high-affinity RAN-GTP-binding proteins, RANBP1 and RANBP2 (or NUP358). This enables conversion of RAN-GTP to RAN-GDP and binding of importin-β1 to new substrates.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Principles of Virology»
Представляем Вашему вниманию похожие книги на «Principles of Virology» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Principles of Virology» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.