Jane Flint - Principles of Virology
Здесь есть возможность читать онлайн «Jane Flint - Principles of Virology» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.
- Название:Principles of Virology
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Principles of Virology: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Principles of Virology»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Volume I: Molecular Biology
Volume II: Pathogenesis and Control
Principles of Virology, Fifth Edition
Principles of Virology — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Principles of Virology», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Viruses are not just for infections anymore! They will provide a rich source of building blocks for applications spanning the worlds of molecular biology, materials science, and medicine.
Lee BY, Zhang J, Zueger C, Chung WJ, Yoo SY, Wang E, Meyer J, Ramesh R, Lee SW. 2012. Virus-based piezoelectric energy generation. Nat Nanotechnol 7:351–356.
Nam KT, Kim DW, Yoo PJ, Chiang CY, Meethong N, Hammond PT, Chiang YM, Belcher AM. 2006. Virus-enabled synthesis and assembly of nanowires for lithium ion battery electrodes. Science 312:885–888.
Tarascon JM. 2009. Nanomaterials: viruses electrify battery research. Nat Nanotechnol 4:341–342.
Wang Q, Lin T, Tang L, Johnson JE, Finn MG. 2002. Icosahedral virus particles as addressable nanoscale building blocks. Angew Chem Int Ed Engl 41:459–462.
The internal components of influenza A virus particles differ radically: they comprise not a single nucleocapsid but multiple ribonucleoproteins, one for each of the 8 molecules of the segmented RNA genome present in an infectious virus particle ( Appendix, Fig. 15). Furthermore, with the exception of terminal sequences, the RNA in these ribonucleoproteins is fully accessible to solvent, suggesting that the RNA is not sequestered in the interior of the ribonucleoprotein. The architectures of ribonucleoproteins released from influenza A virus particles determined by cryo-EM or scanning transmission EM tomography are consistent with such a model: the ribonucleoprotein comprises a double helix of NP molecules connected at one end by an NP loop, often with a molecule of the viral RNA polymerase bound at the other end ( Fig. 4.8A). The RNA is bound along the exposed surfaces of the NP strands with some sequences in each RNA segment more tightly associated than others ( Fig. 4.8B).
The examples presented above illustrate the diversity possible when viruses with simple helical symmetry possess an envelope. Exceptionally large examples include the (+) strand RNA virus potato virus Y, up to 900 nm in length, and bacterial inoviruses, some twice as long, that contain single-stranded DNA genomes. Nevertheless, helical viruses are limited in size. Because helical structures are “open,” some property other than symmetry must limit the size of helical viruses, perhaps the nature of their genomes (see Chapter 3) or susceptibility to shear forces.
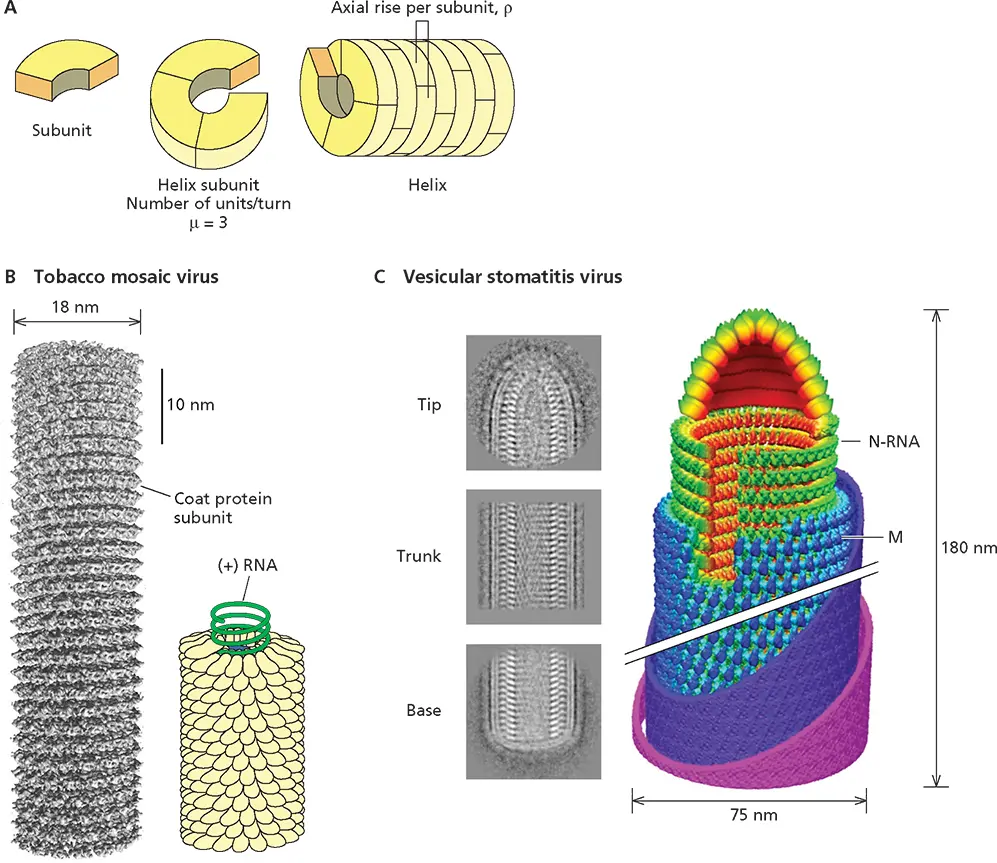
Figure 4.6 Virus structures with helical symmetry. (A)Schematic illustration of a helical particle, indicating the individual subunits, their interaction to form a helical turn, the helix, and the helical parameters ρ (axial rise per subunit) and μ (the number of subunits per turn). The pitch of the helix, P , is given by the formula P = ρ × μ. (B)Tobacco mosaic virus. (Left) A cryo-EM reconstruction at <5-Å resolution of a 70-nm segment of this particle. Each helical turn contains 16.3 protein molecules. Reprinted from Sachse J et al. 2007. J Mol Biol 371:812–835, with permission. Courtesy of N. Grigorieff, Leibniz-Institut für Alterforschung, Jena, Germany. (Right) The regular interaction of the (+) strand RNA genome with coat protein subunits is illustrated in the model based on an X-ray diffraction structure. Data from Namba K et al. 1989. J Mol Biol 208:307–325. (C) Vesicular stomatitis virus. Representative averages of cryo-EM images of the central trunk, conical tip, and flat base of this bullet-shaped virus particle are shown at the left. The trunk and tip were analyzed and reconstructed separately to form the montage model shown on the right, with N and M proteins in green and blue, respectively, and the membrane in purple and pink. The N protein packages the (−) strand RNA genome in a left-handed helix. The crystal structure of N determined in an N-RNA complex ( Fig. 4.7) fits unambiguously with the cryo-EM density of trunk N subunits. The turns of the N protein helix are not closely associated with one another, a property that accounts for the unwinding of the nucleoprotein in the absence of M (see text), which forms an outer, left-handed helix. At the tip, N molecules interact in the absence of RNA. In the trunk, the N helix contains 37.5 subunits per turn. Comparison of N-N interactions in such a turn and in rings of 10 N molecules ( Fig 4.7), as well as the results of mutational analysis, are consistent with formation of rings containing increasing numbers of N molecules from the tip via different modes of N-N interaction induced by association with long genomic RNA. Once a second turn of the N-RNA is stacked on the first, the M protein can bind to add rigidity. Reprinted from Ge P et al. 2010. Science 327:689–693, with permission. Courtesy of Z.H. Zhou, University of California, Los Angeles.
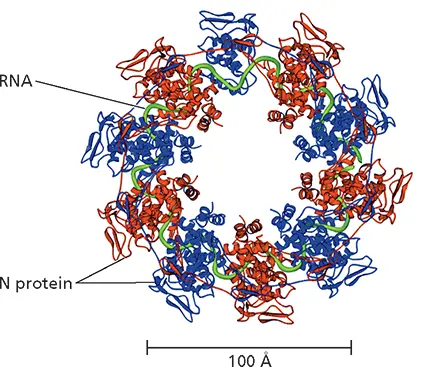
Figure 4.7 Structure of a ribonucleoprotein-like complex of vesicular stomatitis virus. Shown is the structure of a decamer of the N protein bound to RNA, determined by X-ray crystallography, with alternating monomers in the ring colored red and blue and the RNA ribose-phosphate backbone depicted as a green tube. To allow visualization of the RNA, the C-terminal domain of the monomer at the top center is not shown. The decamer was isolated by dissociation of the viral P protein from RNA-bound oligomers formed when the N and P proteins were synthesized in Escherichia coli . Although considerably smaller than N-RNA rings in the virus particles, this structure revealed how N protein molecules interact with the RNA genome and with one another. For example, the N-terminal extension and the extended loop in the C-terminal lobe contribute to the extensive interactions among neighboring N monomers. Adapted from Green TJ et al. 2006. Science 313:357–360, with permission. Courtesy of M. Luo, University of Alabama at Birmingham.
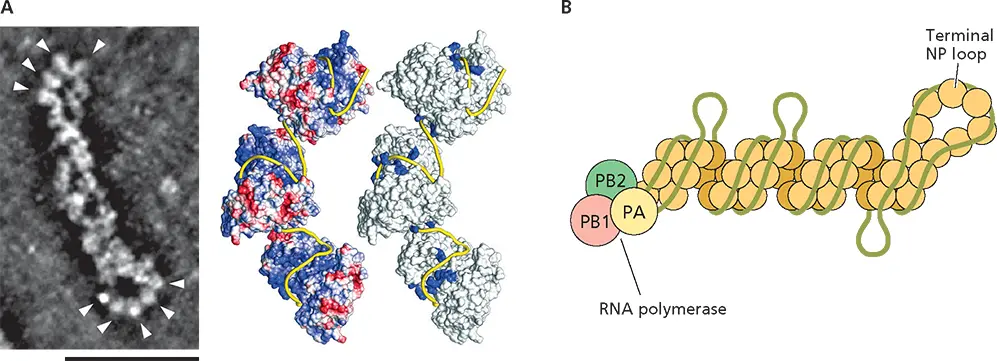
Figure 4.8 Structure of an influenza A virus ribonucleoprotein. (A)(Left) Ribonucleoproteins (RNPs) were isolated from purified influenza A virus particles and examined by scanning transmission EM tomography. Shown is a single RNP segment, with the NP loops indicated by arrowheads: most RNPs have the viral RNA polymerase bound at the end opposite the NP loop. Scale bar, 50 nm. Adapted from Sugita Y et al. 2013. J Virol 87:12879–12884. Courtesy of Y. Kawoaka, University of Tokyo, Japan. (Right) Central and terminal regions of purified RNPs were analyzed separately following cryo-EM. This procedure was adopted to overcome the heterogeneity in length of individual RNPs and their flexibility. Class averaging of images of straight segments of central regions and three-dimensional reconstruction revealed that the RNA-binding NP protein forms a double helix closed by a loop at one end. The likely localization of the (−) strand genome RNA (yellow ribbon) was deduced from the surface electrostatic potential (left, with positive and negative charge shown in blue and red, respectively) and the positions of substitutions that impair binding of NP to RNA (blue in the model on the right). Reprinted from Arranz R et al. 2012. Science 338:1634–1637, with permission. Courtesy of J. Martin-Benito, Centro Nacional de Biotecnologia, Madrid, Spain. (B)Non-uniform association of (–) strand RNA segments with the NP double helix is illustrated schematically, with the NP strands of opposite polarity shown in pale and dark tan; the RNA polymerase subunits at the other end in green, pink, and yellow; and the RNA shown in green. This mode of association, in which G-rich sequences in each RNA genome segment are more tightly bound, was deduced from high-throughput sequencing of RNA fragments bound to RNPs isolated by immunoprecipitation following UV cross-linking of influenza A virus particles and limited RNase digestion of viral lysates. Adapted from Lee N et al. 2017. Nucleic Acids Res 45:8968–8977, with permission.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Principles of Virology»
Представляем Вашему вниманию похожие книги на «Principles of Virology» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Principles of Virology» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.