S. Jane Flint - Principles of Virology, Volume 2
Здесь есть возможность читать онлайн «S. Jane Flint - Principles of Virology, Volume 2» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Principles of Virology, Volume 2
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Principles of Virology, Volume 2: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Principles of Virology, Volume 2»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Volume I: Molecular Biology
Volume II: Pathogenesis and Control
Principles of Virology, Fifth Edition
Principles of Virology, Volume 2 — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Principles of Virology, Volume 2», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
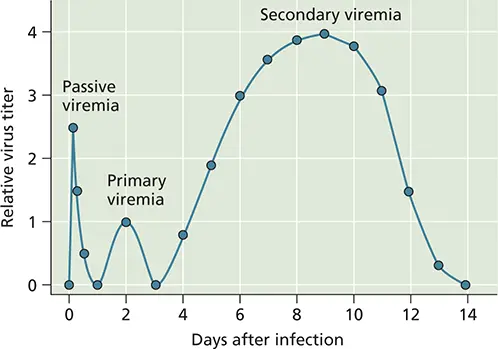
Figure 2.15 Generic characteristics of viremia. Passive viremia occurs when the host is the recipient of infectious virus from an exogenous source (e.g., an infected mosquito). Soon thereafter, a modest primary viremia can occur as a result of virus reproduction at the site of entry. Virus then can be detected in blood, perfusing tissues such as muscle, spleen, and blood vessels. Following reproduction in these sites, a much more robust infection can be detected in the blood, which then can lead to infection of susceptible cells in other organs. Adapted from Nathanson N (ed). 2007. Viral Pathogenesis and Immunity (Academic Press, London, United Kingdom), with permission.
The concentration of virus particles in blood is determined by the rate of their synthesis in permissive tissues and by how quickly they are released into, and removed from, the blood. Circulating particles are engulfed and destroyed by phagocytic cells of the reticuloendothelial systemin the liver, lungs, spleen, and lymph nodes. When serum antibodies appear, virus particles in the blood may be bound by them and neutralized ( Chapter 4). Formation of a complex of antibodies and virus particles facilitates uptake by Fc receptors carried by macrophages lining the circulatory vessels. These virus-antibody complexes can be sequestered in significant quantities in the kidneys, spleen, and liver, prior to elimination from the host via urine or feces. On average, individual virus particles may remain in the blood from 1 to 60 min, depending on parameters such as the physiology of the host (e.g., age and health) and the size and structural integrity of the virus particles. Some viral infections are noteworthy for the long-lasting presence of infectious particles in the blood. Humans infected with hepatitis B and C viruses or mice infected with lymphocytic choriomeningitis virus may have active viremia that persists for months to years. In some cases, movement to the kidney and liver is aided by engagement of virus particles by scavenger receptors found on circulating and resident macrophages. Such receptors bind to common ligands on pathogens, such as lipoproteins, apoptosing cells, cholesterol, and carbohydrates, removing them from the blood. For example, the resident macrophages of the liver, kupffer cells, express high levels of the scavenger receptor type A, which binds to adenovirus particles, targeting them for degradation and elimination from the infected host.
Viremia is of diagnostic value to monitor the course of infection in an individual over time, and epidemiologists use the detection of viremia to identify infected individuals within a population. Frequently, it may be difficult, or technically impossible, to quantify infectious particles in the blood, as is the case for hepatitis B virus. In these situations, the presence of characteristic viral proteins, such as the reverse transcriptase for human immunodeficiency virus type 1, and the presence of the viral genome provide surrogate markers for viremia.
However, the presence of infectious virus particles in the blood also presents practical problems. Infections can be spread inadvertently in the population when pooled blood from thousands of individuals is used for therapeutic purposes (transfusions) or as a source of therapeutic proteins (gamma globulin or blood-clotting factors). We have learned from unfortunate experience that bloodborne viruses, such as hepatitis viruses and human immunodeficiency virus type 1, can be spread by contaminated blood and blood products. The World Health Organization estimates that, as of 2000, inadequate blood screening resulted in 1 million new human immunodeficiency virus type 1 infections worldwide . Careful screening for these viruses in blood supplies before transfusion into patients is now standard procedure. However, sensitive detection methods and stringent purification protocols are useful only when we know what we are looking for; as-yet-undiscovered viruses may still be transmitted through the blood supply ( Box 2.9).
BOX 2.9
TERMINOLOGY
The viruses in your blood
If you have ever received a blood transfusion, along with the red blood cells, leukocytes, plasma, and other components, you also were likely infused with a collection of viruses. A recent study of the blood virome of more than 8,000 healthy individuals revealed a total of 19 different DNA viruses in 42% of the subjects.
Viral DNA sequences were identified among the genome sequences of 8,240 individuals that were determined from blood. Of the 1 petabyte (1 million gigabytes) of sequence data that were generated, ∼5% did not correspond to human DNA. Within this fraction, sequences of 94 different viruses were identified. Nineteen of these were human viruses. The method is not expected to reveal RNA viruses except retroviruses that are integrated as DNA copies in the host chromosomes.
The most common human viruses identified were herpesviruses, including cytomegalo virus, EpsteinBarr virus, herpes simplex virus, and human herpes viruses 7 and 8, found in 14 to 20% of individuals. Anelloviruses, small viruses with a circular genome, were found in 9% of the samples. Other viruses found in less than 1% of the samples included papillomaviruses, parvoviruses, polyomavirus, adenovirus, human immunodeficiency virus type 1, and human Tcell lymphotropic virus (the latter two integrated into host DNA).
The other 75 viruses are likely contaminants from laboratory reagents or the environment. These include sequences from nonhuman retroviruses, four different giant DNA viruses, and a virus of bees, all found in fewer than 10 samples. These findings illustrate the challenge in distinguishing bona fide human viruses from contaminants.
Identifying viruses in blood is an important objective for ensuring the safety of the blood supply. Donor blood is currently screened for human immunodeficiency virus types 1 and 2, human T-cell lymphotropic virus-1 and -2, hepatitis C virus, hepatitis B virus, West Nile virus, and Zika virus. These viruses are pathogenic for humans and can be transmitted via the blood. Some viruses, such as anelloviruses and pegiviruses, are in most donated blood, yet their pathogenic potential is unknown. It is not feasible to reject donor blood that contains any type of viral nucleic acid—if we did, we would not have a blood supply.
Continuing studies of the blood virome are needed to define which viruses should be tested for in donated blood. The human papillomavirus (17 people), Merkel cell polyomavirus (49 people), human herpesvirus type 8 (3 people), and adenovirus (9 people) detected in this study could be transmitted in the blood, and their presence should be monitored in future studies.
It is important to emphasize that this work describes only viral DNA sequences, and not infectious virus particles. The blood supply is screened by nucleic acid tests, but it is crucial to determine if infectious virus particles are also present . If viral DNA is present in blood but particles are never found, then it might not be necessary to reject blood based on the presence of certain sequences.
Moustafa A, Xie C, Kirkness E, Biggs W, Wong E, Turpaz Y, Bloom K, Delwart E, Nelson KE, Venter JC, Telenti A. 2017. The blood DNA virome in 8,000 humans. PLoS Pathog 13:e1006292.
Neural Spread
Some viruses spread from the primary site of infection by entering local nerve endings. In some cases, neuronal spread is the definitive characteristic of pathogenesis, notably by rabies virus and alphaherpesviruses, which cause infections that primarily impact neuronal function or survival. In other cases, invasion of the nervous system is a rare, typically dead-end, diversion from the normal site of reproduction (e.g., poliovirus, reovirus). Mumps virus, rubella virus, human immunodeficiency virus type 1, and measles virus can reproduce in the brain but access the central nervous system by the hematogenous route, often ferried into the brain by infected lymphocytes or monocytes. The molecular mechanisms that dictate spread into the brain by neural or hematogenous pathways are not well understood, and the way these viruses are defined can lead to further confusion ( Box 2.10). For those neurotropic viruses that enter the brain via neuronal circuitry, viral reproduction usually occurs first in nonneuronal cells such as muscle cells near the site of infection. Following reproduction in these cells, virus particles subsequently spread into afferent (e.g., sensory) or efferent (e.g., motor) nerve fibers that innervate the infected tissue, usually crossing neuromuscular junctions to do so ( Fig. 2.16).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Principles of Virology, Volume 2»
Представляем Вашему вниманию похожие книги на «Principles of Virology, Volume 2» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.



Обсуждение, отзывы о книге «Principles of Virology, Volume 2» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.