S. Jane Flint - Principles of Virology, Volume 2
Здесь есть возможность читать онлайн «S. Jane Flint - Principles of Virology, Volume 2» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Principles of Virology, Volume 2
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Principles of Virology, Volume 2: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Principles of Virology, Volume 2»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Volume I: Molecular Biology
Volume II: Pathogenesis and Control
Principles of Virology, Fifth Edition
Principles of Virology, Volume 2 — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Principles of Virology, Volume 2», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Sequences in viral genomes that control transcription of viral genes, such as enhancers, may be determinants of viral tropism. In the brain, JC polyomavirus reproduces only in oligodendrocytes, because the JC virus enhancer is active only in this cell type. Other examples include the liver-specific enhancers of hepatitis B virus and the keratinocyte-specific enhancer of human papillomavirus type 11.
Cellular proteases, present in some cells but not others, are often required to cleave viral proteins to form the mature infectious virus particle (Volume I, Chapter 13). For example, a cellular protease in the lung cleaves the influenza virus HA0 precursor into two subunits so that fusion of the viral envelope and cell membrane can proceed. In mammals, the reproduction of influenza virus is restricted to epithelial cells of the upper and lower respiratory tracts, and its tropism is thought to be influenced by the production of the protease that processes HA0. This serine protease, called tryptase, is secreted by nonciliated club cells of the bronchial and bronchiolar epithelia ( Fig. 2.11), one of the only cell types to do so. Purified tryptase can cleave and activate HA0 in virus particles in vitro . Alteration of the hemagglutinin (HA) cleavage site so that it can be recognized by other cellular proteases dramatically changes the tropism of the virus and its pathogenicity; some highly virulent avian influenza virus strains contain an insertion of multiple basic amino acids at the cleavage site of HA0. This new sequence permits processing by ubiquitous intracellular proteases, such as furins. As a result, these variant viruses are released in active form and are able to infect many organs of birds, including the spleen, liver, lungs, kidneys, and brain. Naturally occurring mutants of this type cause high mortality in poultry farms. These viruses can also infect humans: avian influenza viruses isolated from 16 people in Hong Kong in 1997 contained similar amino acid substitutions at the HA0 cleavage site, and many of these individuals had gastrointestinal, hepatic, and renal symptoms as well as respiratory disease. A virus with such an HA0 site alteration had not been previously identified in humans, and its isolation led to fears that an influenza pandemic was imminent, resulting in the preventative slaughter of all chickens in Hong Kong. Beyond these direct alterations in the viral HA0, changes in other viral proteins can influence HA cleavage indirectly ( Box 2.7).
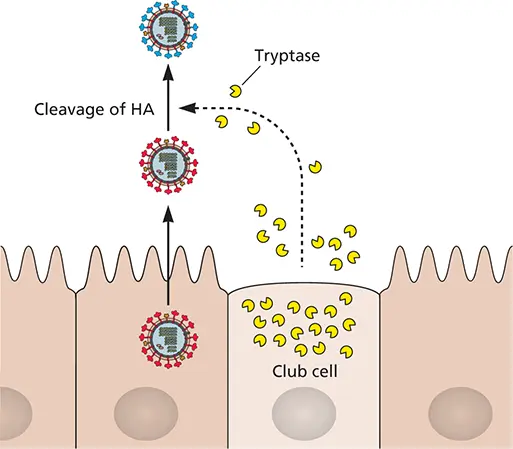
Figure 2.11 Cleavage of influenza virus HA0 by club cell tryptase. Influenza viruses reproduce in respiratory epithelial cells in humans. These virus particles contain the uncleaved form of HA (HA0) and are noninfectious. Club cells secrete a protease, tryptase, which cleaves the HA0 of extracellular particles, thereby rendering the viral particles infectious. Adapted from Tashiro M, Rott R. 1996. Semin Virol 7:237–243, with permission. Note: In previous editions of this text, club cells were referred to as “Clara cells,” named after the German scientist who discovered them. Because Clara was an active member of the Nazi party, in 2013, the lung physiology community elected to change the name of these cells to “club cells.” We have adopted this convention.
BOX 2.7
DISCUSSION
A mechanism for expanding the tropism of influenza virus is revealed by analyzing infections that occurred in 1940
Entry of influenza virus is controlled by two glycoproteins, hemagglutinin (HA) and neuraminidase (NA), present on the viral surface. Initiation of virus infection involves binding to sialic acids on carbohydrate side chains of cellular glycoproteins and glycolipids. Until the isolation of the H5N1 virus from 16 individuals in Hong Kong, viruses with the HA0 cleavage site mutation that permits cleavage by ubiquitous furin proteases had not been found in humans. Similarly, the WSN/33 strain of influenza virus, produced in 1940 by passage of a human isolate in mouse brain, is pantropic in mice. Unlike most human influenza virus strains, WSN/33 can reproduce in cells in culture in the absence of added trypsin, because its HA0 can be cleaved by serum plasmin. Surprisingly, it was found that the NA of WSN/33 is necessary for HA0 cleavage by serum plasmin. This altered NA protein can bind plasminogen, sequestering it on the cell surface, where it is converted to the active form, plasmin (see figure, panel A). Plasmin then cleaves HA0 into HA1 and HA2. Therefore, a change in NA, not in HA, allowed cleavage of HA by a ubiquitous cellular protease. This property may, in part, explain the pantropic nature of WSN/33.
Goto H, Kawaoka Y. 1998. A novel mechanism for the acquisition of virulence by a human influenza A virus. Proc Natl Acad Sci U S A 95:10224–10228.
Taubenberger JK. 1998. Influenza virus hemagglutinin cleavage into HA1, HA2: no laughing matter. Proc Natl Acad Sci U S A 95:9713–9715.
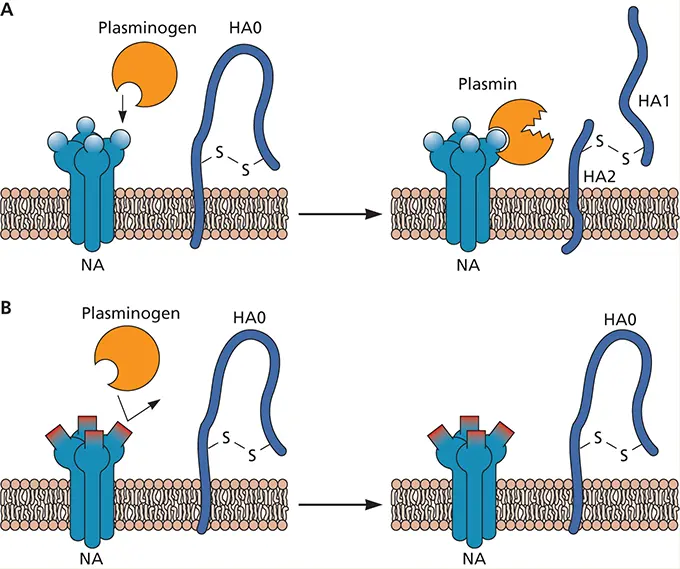
Proposed mechanism for activation of plasminogen and cleavage of HA. (A)Plasminogen binds to NA, which has a lysine at the carboxyl terminus. A cellular protein converts plasminogen to the active form, plasmin. Plasmin then cleaves HA0 into HA1 and HA2. (B)When NA does not contain a lysine at the carboxyl terminus, plasminogen cannot interact with NA and is not activated to plasmin. Therefore, HA is not cleaved. Data from Goto H, Kawaoka Y. 1998. Proc Natl Acad Sci U S A 95:10224–10228.
A final emerging class of cellular macromolecules that can influence viral tropism are small, non-protein-encoding RNA species, called microRNAs. Although these RNAs do not result in new protein production, they can still dramatically affect host cell physiology. For example, microRNA-122 is conserved among vertebrates (but not present in invertebrates), and its expression is highest in the liver, where it likely contributes to fatty acid metabolism. The liver-tropic virus hepatitis C virus depends on microRNA-122 for reproduction. This microRNA binds directly to two adjacent sites close to the 5′ end of hepatitis C virus RNA, impacting RNA stability and genome replication (Volume 1, Chapter 9).
As we learn more about viruses, hosts, and populations, we are discovering that some surprising variables, including the gender of the host, can influence both the frequency and severity of viral infections ( Box 2.8).
Spread throughout the Host
Following reproduction at the site of entry, virus particles can remain localized or can spread to other tissues. Spread beyond the initial site of infection depends on multiple parameters, including the initial viral dose, the presence of viral receptors on other cells, and the relative rates of immune induction and release of infectious virus particles. Localized infections in the epithelium are usually limited by the physical constraints of the tissue and are brought under control by the intrinsic and innate immune defenses discussed in Chapter 3. An infection that spreads beyond the primary site (usually near the point of viral entry) is said to be disseminated. If many organs are viral targets, the infection is described as systemic. Spread beyond the primary site requires continued breaching of the host’s physical barriers. For example, virus particles may be able to cross a basement membrane when the integrity of that structure is compromised by inflammation and epithelial cell destruction. Below the basement membrane are subepithelial tissues, where virus particles encounter tissue fluids, the lymphatic system, and phagocytes. These host components make substantial contributions to clearing foreign particles, but may also allow infectious virus particles to be carried beyond the primary site of infection.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Principles of Virology, Volume 2»
Представляем Вашему вниманию похожие книги на «Principles of Virology, Volume 2» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.



Обсуждение, отзывы о книге «Principles of Virology, Volume 2» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.