S. Jane Flint - Principles of Virology, Volume 2
Здесь есть возможность читать онлайн «S. Jane Flint - Principles of Virology, Volume 2» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Principles of Virology, Volume 2
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Principles of Virology, Volume 2: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Principles of Virology, Volume 2»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Volume I: Molecular Biology
Volume II: Pathogenesis and Control
Principles of Virology, Fifth Edition
Principles of Virology, Volume 2 — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Principles of Virology, Volume 2», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Neurons are polarized cells with structurally and functionally distinct processes (axons and dendrites) that can be separated by enormous distances. For example, in adult humans, the axon terminals of motor neurons that control stomach muscles can be 50 centimeters away from the cell bodies and dendrites in the brain stem. Certainly, neurotropic viruses do not traverse these great distances by Brownian (random) motion. Rather, the neuronal cytoskeleton, including microtubules and actin, provides the “train tracks” that enable movement of mitochondria, synaptic vesicles, and virus particles to and from the synapse. Molecular motor proteins, such as dyneins and kinesins, are the “engines” that move along these cellular highways ( Box 2.11). Drugs, such as colchicine, that disrupt microtubules efficiently block the spread of many neurotropic viruses from the site of peripheral inoculation to the central nervous system.
With few exceptions, cells of the peripheral nervous system are the first to be infected by neurotropic viruses. These neurons represent the first cells in circuits connecting the innervated peripheral tissue with the spinal cord and brain. Once in the nervous system, alphaherpesviruses and some rhabdoviruses (e.g., rabies virus), flaviviruses (e.g., West Nile virus), and paramyxoviruses (e.g., measles and canine distemper virus) can spread among neurons connected by synapses ( Box 2.11). Virus spread by this mode can continue through chains of connected neurons of the peripheral nervous system and may eventually reach the spinal cord and brain, often with devastating results ( Fig. 2.17). Nonneuronal support cells and satellite cells in ganglia may also become infected.
Movement of virus particles and their release from infected cells are important features of neuronal infections. As is true for polarized epithelial cells discussed earlier, directional release of virus particles from neurons affects the outcome of infection. Alphaherpesviruses become latent in peripheral neurons that innervate the site of infection. Reactivation from the latent state results in viral reproduction in the primary neuron and subsequent transport of progeny virus particles from the neuron cell body back to the innervated peripheral tissue where the infection originated. Alternatively, virus particles can spread from the peripheral to the central nervous system. The direction taken matters tremendously: going one way results in a minor local infection (a cold sore); going the other way can cause a life-threatening viral encephalitis. Luckily, spread back to the peripheral site (away from the brain) is far more common.
BOX 2.10
TERMINOLOGY
Infection of the nervous system: definitions and distinctions
A neuroinvasive virus can enter the central nervous system (spinal cord and brain) after infection of a peripheral site.
A neurotropic virus can infect neurons; infection may occur by neural or hematogenous spread from a peripheral site.
A neurovirulent virus can cause disease of nervous tissue, manifested by neurological symptoms and often death.
Examples:
Herpes simplex virus type 1 exhibits low neuroinvasiveness but high neuroviru lence. It always enters the peripheral nervous system but rarely gains access to the central nervous system. When it does, the consequences are severe, and can be fatal. Mumps virus exhibits high neuroinvasiveness but low neurovirulence. Most infections lead to invasion of the central nervous system, but neurological disease is mild. Rabies virus is highly neuroinvasive, with high neurovirulence. It readily infects the peripheral nervous system and spreads to the central nervous system with 100% lethality, unless postinfection vaccination is given.
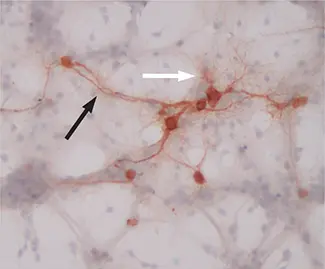
Primary mouse hippocampal neurons expressing a measles virus receptor, CD46, and infected with measles virus for 48 h.Virus-infected cells are stained brown. Black arrow: neuronal axon; white arrow: neuronal dendrites. Original magnification = ×200. Photo courtesy of the Rall laboratory.
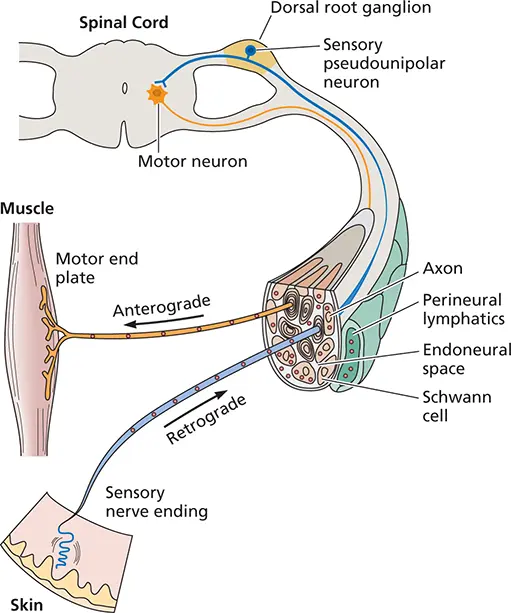
Figure 2.16 Possible pathways for the spread of infection in nerves. Virus particles may enter sensory or motor neuron endings. They may be transported within axons, in which case viruses taken up at sensory endings reach dorsal root ganglion cells. Those taken up at motor endings reach motor neurons. Viruses may also travel in the endoneural space, perineural lymphatics, or infected Schwann cells. Directional transport of virus particles inside the sensory neuron is defined as anterograde [movement from the (–) to the (+) ends of microtubules] or retrograde (vice versa).
Organ Invasion
Once virus particles enter the blood or neurons and are dispersed from the primary site of infection, any subsequent reproduction requires invasion of other cells. We have already discussed viral movement into and among neurons to access the brain and spinal cord and will return to this issue in Chapter 5when we discuss neuropathogenesis resulting from viral infections.
There are three main types of blood vessel-tissue junctions that serve as portals for tissue invasion ( Fig. 2.18). In some tissues, the endothelial cells are continuous with a dense basement membrane. At other sites, the endothelium contains gaps, and at still others, there may be sinusoids, in which macrophages form part of the blood-tissue junction. Viruses can traverse all three types of junctions.
Entry into Organs with Sinusoids
Organs such as the liver, spleen, bone marrow, and adrenal glands are characterized by the presence of sinusoids, a type of capillary that has a large pore size to allow free exchange between the blood and the tissue. These capillaries are lined with macrophages, known somewhat misleadingly as the reticuloendothelial system (these macrophages are neither endothelial nor a “system”). These cells function to filter the blood and remove foreign particles, similar to a HEPA filter purifying incoming air. However, the macrophages in organs that possess sinusoids often provide the portal for entry of viral particles into tissues. For example, hepatitis viruses that infect the liver, the major filtering and detoxifying organ of the body, usually enter from the blood. The presence of virus particles in the blood can lead to the infection of Kupffer cells, the macrophages that line liver sinusoids ( Fig. 2.19). Virus particles may be transcytosed across kupffer and endothelial cells without reproduction to reach the underlying hepatocytes. Alternatively, viruses may multiply in the kupffer cells, and then be released to infect hepatocytes. Either mechanism may induce inflammation and necrosis of liver tissue, a condition termed hepatitis.
Entry into Organs That Lack Sinusoids
To enter tissues that lack sinusoids ( Fig. 2.18), and that have tighter or more continuous capillary endothelial cells, virus particles must first adhere to the endothelial cells before crossing into the tissue. This often occurs in venules, where the flow is slowest and the capillary walls are thinnest. High viral loads and persistence of viral particles in the blood stream enhance the likelihood of tissue penetration. Once blood-borne virus particles have adhered to the vessel wall, they can more easily invade the renal glomerulus, pancreas, ileum, or colon. Invasion occurs because the endothelial cells that make up the capillaries of these tissues have pores or holes in the cell layer, called fenestrations, that permit virus particles or virus-infected cells to cross. Some viruses traverse the endothelium hidden in infected monocytes or lymphocytes, a process called diapedesis.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Principles of Virology, Volume 2»
Представляем Вашему вниманию похожие книги на «Principles of Virology, Volume 2» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.



Обсуждение, отзывы о книге «Principles of Virology, Volume 2» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.