Michael Begon - Ecology
Здесь есть возможность читать онлайн «Michael Begon - Ecology» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Ecology
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Ecology: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Ecology»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
– now in full colour – offers students and practitioners a review of the ecological sciences.
The previous editions of this book earned the authors the prestigious ‘Exceptional Life-time Achievement Award’ of the British Ecological Society – the aim for the fifth edition is not only to maintain standards but indeed to enhance its coverage of Ecology.
In the first edition, 34 years ago, it seemed acceptable for ecologists to hold a comfortable, objective, not to say aloof position, from which the ecological communities around us were simply material for which we sought a scientific understanding. Now, we must accept the immediacy of the many environmental problems that threaten us and the responsibility of ecologists to play their full part in addressing these problems. This fifth edition addresses this challenge, with several chapters devoted entirely to applied topics, and examples of how ecological principles have been applied to problems facing us highlighted throughout the remaining nineteen chapters.
Nonetheless, the authors remain wedded to the belief that environmental action can only ever be as sound as the ecological principles on which it is based. Hence, while trying harder than ever to help improve preparedness for addressing the environmental problems of the years ahead, the book remains, in its essence, an exposition of the
of ecology. This new edition incorporates the results from more than a thousand recent studies into a fully up-to-date text.
Written for students of ecology, researchers and practitioners, the fifth edition of
is anessential reference to all aspects of ecology and addresses environmental problems of the future.
Ecology — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Ecology», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Source : (a, b) After de Wit (1965) and other sources. (c) After Meshram et al . (2017).
shade: resource‐depletion zones and spectral changes
Less systematic variations in the radiation environment of a leaf are caused by the nature and position of neighbouring leaves. Leaves in a canopy, by intercepting radiation, create a resource‐depletion zone (RDZ) – in this case, a moving band of shadow over other leaves of the same plant, or of others. The composition of radiation that has passed through leaves in a canopy, or through a body of water, is also altered. Typically, it is depleted in the blue and (especially through water) the red parts of the spectrum – the most effective wavelengths for photosynthesis. Figure 3.3shows an example for the variation with depth in a freshwater habitat.

Figure 3.3 The spectral distribution of radiation changes with depthas shown here for Lake Burley Griffin, Australia. Note that photosynthetically active radiation lies broadly within the range 400–700 nm.
Source : After Kirk (1994).
sun and shade species
The way in which organisms react to systematic, predictable patterns in the supply of a resource reflects both their present physiology and their past evolution. At a very broad scale, the seasonal shedding of leaves by deciduous trees in temperate regions in part reflects the annual rhythm in the intensity of radiation – they are shed when they are least useful. Amongst terrestrial species, plants that are characteristic of shaded habitats generally use radiation at low intensities more efficiently than sun species, but the reverse is true at high intensities ( Figure 3.4). Part of the difference between them lies in the physiology of the leaves, but the morphology of the plants also influences the efficiency with which radiation is captured. The leaves of sun plants are commonly exposed at acute angles to the midday sun, spreading an incident beam of radiation over a larger leaf area and effectively reducing its intensity (Poulson & DeLucia, 1993). The leaves of sun plants are also usually superimposed into a multilayered canopy. In bright sunshine even the shaded leaves in lower layers may have positive rates of net photosynthesis. Shade plants adopt a different strategy, commonly having leaves held near to the horizontal and in a single‐layered canopy.
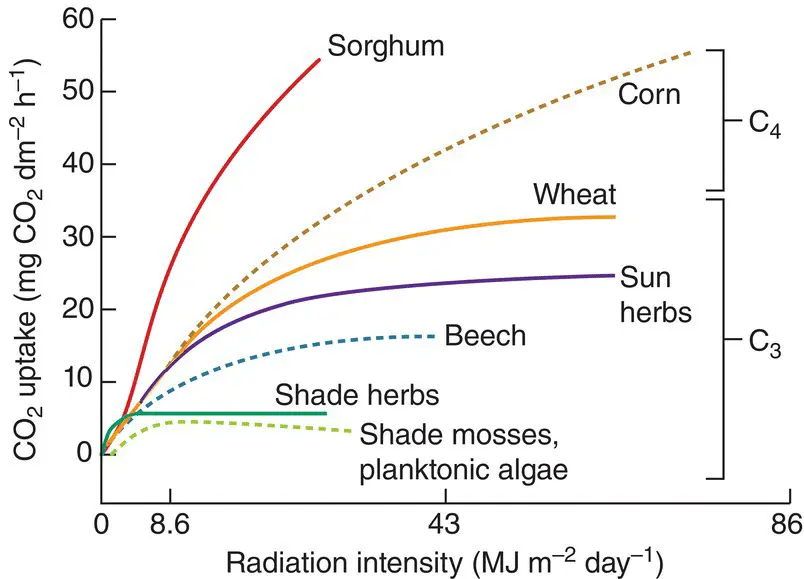
Figure 3.4 The response of photosynthesis to radiation intensity in various plants at optimal temperatures and with a natural supply of CO 2. Note that corn and sorghum are C 4plants and the remainder are C 3(the terms are explained in Sections 3.3.1and 3.3.2).
Source : After Larcher (1980), and other sources.
sun and shade leaves
Plants may also respond ‘tactically’ to the radiation environment in which they develop, producing ‘sun leaves’ and ‘shade leaves’ within the canopy of a single plant. Sun leaves (and indeed, leaves on sun plants) are typically smaller, thicker, have more cells per unit area, denser veins, more densely packed chloroplasts and a greater dry weight per unit area of leaf. They are said to have a smaller specific leaf area (leaf area per unit leaf mass). Acclimation to shade typically involves increasing chlorophyll concentration and decreasing investment in the rest of the photosynthetic apparatus. This allows the leaf to maximise capture of light, but does not waste resource on a high photosynthetic capacity, which is not needed under shade conditions. In turn, this releases nitrogen for use by the upper leaves. However, these tactical manoeuvres take time. It is impossible for the plant to change its form fast enough to track the changes in intensity of radiation between a cloudy and a clear day. It can, however, change its rate of photosynthesis extremely rapidly, reacting even to the passing of a fleck of sunlight.
APPLICATION 3.1 Bioengineering crops for accelerated recovery from photoprotection
The responses of plants to changes in the quantity of light include the induction at high light intensities of photoprotective mechanisms that prevent the photosynthetic machinery from getting ‘overexcited’ and risking the generation of damaging oxidising radicals, instead dissipating excess light as heat. However, when intensities return to harmless levels, there is typically a delay before these protective mechanisms are fully switched off, such that rates of photosynthesis at these times are lower than they might otherwise be. Some calculations suggest that this could cost field crops as much as 20% of their potential yield (Kromdijk et al ., 2016). It would therefore clearly be valuable if that switching off of the protective mechanisms could be speeded up. Bioengineering (the insertion of new or altered genes into a plant) offers the opportunity of applying our understanding of the physiology of photoprotection to effect such an accelerated response. Results are shown in Figure 3.5for a study in which variants of three different genes known to be instrumental in the operation of the mechanism were selected for increased expression levels, following a screen of seedlings of the model plant Arabidopsis thaliana . These variants were then inserted into tobacco plants, Nicotiana tabacum , itself used as a model for crop plants in general, since the photoprotective mechanism being altered is common to all plants.

Figure 3.5 Bioengineering of photoprotection can improve crop plant performance.(a) To the left, a comparison for two measures of photosynthetic efficiency (of CO 2uptake and of electron transport) and of the rate of harmlessly dissipating excess light as heat – the rate of ‘quenching’ of chlorophyll fluorescence (NPQ) – at steady levels of light, between wild type (WT) Arabidopsis plants and three strains bioengineered to switch off photoprotection more rapidly. There were no differences. To the right, a similar comparison but with fluctuating light levels. The bioengineered strains were all significantly more efficient in photosynthesis than the wild type because fluorescence was dampened down more rapidly. (b) The consequences for the bioengineered plants in terms of weight, leaf area and plant height, following 22 days of growth in the field. All strains grew better. In both (a) and (b), bars are SEs and * indicates a significant difference between bioengineered lines and the wild type ( P < 0.05)
Source : After Kromdijk et al . (2016).
When the supply of light was constant, all three types of bioengineered plant behaved similarly to wild type plants in terms of photosynthetic efficiency and the harmless dissipation (‘quenching’) of excess light as heat ( Figure 3.5a, left). But in the field, most leaves experience continually fluctuating light due to clouds and intermittent shading from the leaves above. It is notable, therefore, that in the fluctuating regime, photosynthetic efficiency was higher in the bioengineered plants than the wild types, and their overall level of quenching was lower, because it was compressed into a shorter period ( Figure 3.5a, right). As a result, the bioengineered plants grew much better than the wild types ( Figure 3.5b). Bioengineering of any sort must always be applied with caution, but these results do hold out the prospect of significant increases in yield for a wide variety of crops, since this process is common to all land plants.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Ecology»
Представляем Вашему вниманию похожие книги на «Ecology» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Ecology» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.