Pathology of Genetically Engineered and Other Mutant Mice
Здесь есть возможность читать онлайн «Pathology of Genetically Engineered and Other Mutant Mice» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.
- Название:Pathology of Genetically Engineered and Other Mutant Mice
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Pathology of Genetically Engineered and Other Mutant Mice: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Pathology of Genetically Engineered and Other Mutant Mice»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
An updated and comprehensive reference to pathology in every organ system in genetically modified mice Pathology of Genetically Engineered and Other Mutant Mice
Pathology of Genetically Engineered and Other Mutant Mice
Pathology of Genetically Engineered and Other Mutant Mice — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Pathology of Genetically Engineered and Other Mutant Mice», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
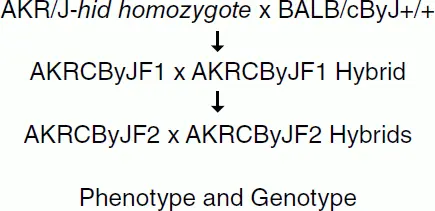
Figure 3.5 Setting up a mapping cross using F1 and F2 hybrids.
Backcrossing an F1 hybrid to one of the parents, instead of sibling intercrossing, is the initial process in the creation of a congenic, consomic, or conplastic strain, and such an N2 population is a useful tool for mapping dominant mutations. Some mutations or transgenes are adequately deleterious to overall health that they cannot be maintained on an inbred background and instead require hybrid vigor. Such mutants are maintained by breeding to an F1 hybrid at each generation or through outcross–intercross to an F1 hybrid. One complicated example of this is B6EiC3Sn a / A ‐Ts(17 16)65Dn/J [11]. This chromosomal aberration was induced in DBA/2J inbred mice, but is detrimental to health and causes male sterility so in order to successfully maintain a colony a female carrier is bred to a (C57BL/6JEiJ x C3H/HeSnJ)F1/J male at each generation. There is no punctuation in the genetic background of the strain name, B6EiC3Sn, and the presence of the DBA/2J contribution is omitted from the nomenclature for simplicity and must be found in sources outside of the strain nomenclature.
Table 3.7 Breeding data used to identify the gene responsible for the hair interior defect phenotype.
| AKR/J‐ hid *homozygote crossed with | Total progeny | Normal mice | Affected mice | Affected (%) | X 2 | p ‐Value |
|---|---|---|---|---|---|---|
| BALB/cByJ | 260 | 222 | 38 | 14.62 | 5.92 | 0.025–0.010 |
| C57BL/6J | 430 | 330 | 100 | 23.26 | 0.220 | 0.500–0.750 |
| CAST/EiJ | 339 | 255 | 84 | 24.78 | 0.003 | >0.900 |
| C3H/HeJ | 248 | 191 | 57 | 22.98 | 0.170 | 0.500–0.750 |
| FVB/NJ | 272 | 205 | 67 | 24.63 | 0.006 | >0.900 |
| Total F2 mice | 1549 | 1203 | 346 |
*AKR/J‐ hid was the original designation until the gene responsible was identified. Because the gene was identified with a different phenotype earlier, the allele is officially Soat1 aldnot Soat1 hid.
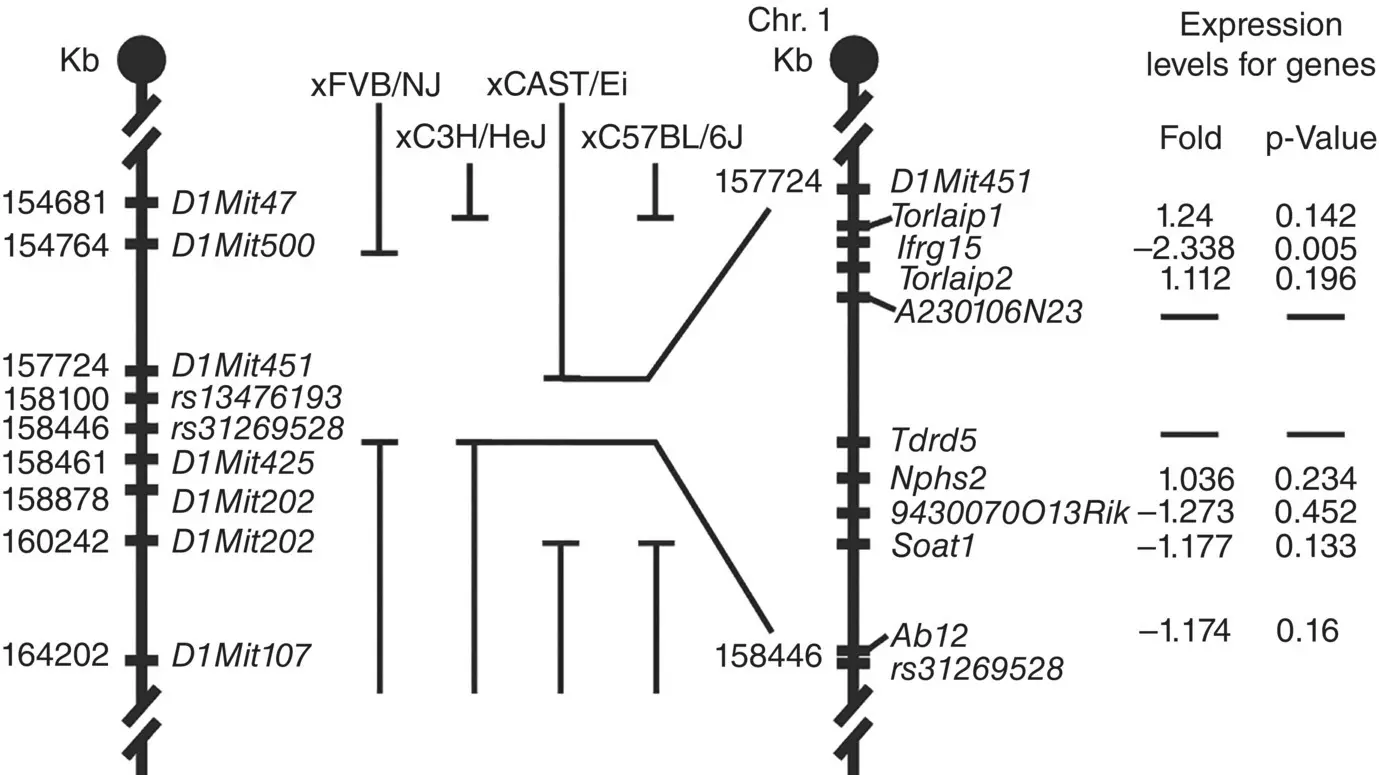
Figure 3.6 Crossing AKR/J mice with more than one inbred strain to map the mutant gene locus. By using the shortest interval between different crosses, the list of candidate genes could be reduced. Sequencing then only had to be done on a few genes to identify the mutant gene responsible for the hair interior defect phenotype.
Sources: Based on Giehl KA et al. [9].
Some mutations are maintained on a background in which the genetic contributions of two inbred strains are segregating. The retarded hair growth (in ornithine aminotransferase, Oat rhg) mutation arose spontaneously in AKR/J and was later outcrossed to C57BL/6JEi and subsequently maintained by sibling intercrossing [12, 13]. Thus, the genetic background consists of contributions from both AKR/J and C57BL/6JEi segregating at each generation. A semicolon between the strain abbreviations is used to represent this situation, B6Ei;AKR‐ Oat rhg/J. If more than two inbred strains contribute to the genetic background of a strain, then it is simply called a stock, such as STOCK Noto tc/J (notochord homeobox gene, truncate mutation). The truncate mutation arose spontaneously in the F4 generation of a cross between a C3H female and a Swiss outbred male. It was subsequently bred to BALB/cHu and again to C3H before being inbred for more than 80 generations [14]. The STOCK is in capital letters designating inbred, and there is no hyphen between STOCK and the allele symbol. The strain could have been assigned a unique inbred strain name, such as TC/J, but was instead designated as a STOCK. When stock is in lower case, it indicates that the segregating population has not reached F20.
Recombinant Inbred (RI) and Recombinant Congenic Mice
One variation of inbred mice is the creation of recombinant inbred (RI) mice. In this case, two different inbred strains are crossed to produce F1 progeny. As with creating inbred strains, breeding pairs of these F1 progeny are crossed to create an F2 population and then one pair is selected for breeding in this and each subsequent generation until at 20 consecutive generations of breeding (F20) from a single brother–sister pair a new inbred strain is created, which has approximately 50% of its DNA derived from each of the original parental inbred strains. The generation of many separate recombinant inbred strains from the same parental inbred strains creates a panel of recombinant inbred stains, each of which has an array of unique DNA recombinations and therefore unique subset of parental DNA segments. Recombinant inbred strains are named with a pair of 1‐ or 2‐letter strain abbreviation codes representing the original parental inbred strains ( Figure 3.3) with the female founder strain first, followed by an X for the cross, then the male founder strain abbreviation followed by a slash, a number to indicate the specific line and then the laboratory code for who made the strain ( Table 3.8). The higher the number of strains in a recombinant inbred panel, the potentially more valuable the collection due to the increased number of recombination sites available for comparison across the panel.
Table 3.8 Recombinant inbred and recombinant congenic strain designation.
| BXD31/TyJ | AcB51/Prgs |
|---|---|
| B= C57BL/6J | A= backcross to A/J |
| X= cross | c= recombinant congenic cross |
| D= DBA/2J | B= C57BL/6J |
| 31= RI line number | 51= recombinant congenic line number |
| Ty= Lab that created the line (Benjamin Taylor) | Prgs= Lab that created the line (Philippe Gros) |
| J= Subsequent breeder of the strain, The Jackson Laboratory |
The Jackson Laboratory Repository distributes more than 125 BXD RI lines and there are many more in private laboratories. Many of these BXD RI lines have been sequenced and annotated. Using websites, such as GeneNetwork ( www.genenetwork.org) [15], it is possible to use these mice for localizing, if not necessarily identifying, the gene(s) responsible for a disease, response to a treatment, or for any phenotype observed in one of the parental strains but not the other. For example, if the primary gene responsible for a disease in DBA/2J mice is known, such as a hypomorphic allele of Abcc6 that causes pseudoxanthoma elasticum, and the gene is wildtype in C57BL/6J, by first identifying all the BXD lines that are homozygous for the D2 mutant allele, one can then select only those mice, phenotype them, and rapidly identify narrow genetic intervals from which to generate a small list of candidate disease modifier genes [16] ( Figure 3.7). Other RI lines exist for a variety of crosses that can be used in a similar manner.
A variation on the RI line is the recombinant congenic strains that are created by crossing two inbred strains followed by one or several backcrosses to one of the parental strains. The resulting mice are then inbred without selection. Recombinant congenic strains are also named using 1‐ or 2‐letter strain abbreviation codes representing the parental strains, but a lower case c for congenic, not an X, separates them and the first strain listed is the host strain, that to which the backcrossing is done, and the strain symbol following the c is the donor strain, that contributing the least amount to the total genetic background. The AcB51/Pgrs recombinant congenic strain was generated by Dr. Philippe Gros by backcrossing (A/J x C57BL/6J)F1 to A/J for two consecutive generations then sibling inbreeding to generate a panel of recombinant consomics including this, line 51.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Pathology of Genetically Engineered and Other Mutant Mice»
Представляем Вашему вниманию похожие книги на «Pathology of Genetically Engineered and Other Mutant Mice» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Pathology of Genetically Engineered and Other Mutant Mice» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.