Genetic Analysis of Complex Disease
Здесь есть возможность читать онлайн «Genetic Analysis of Complex Disease» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Genetic Analysis of Complex Disease
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Genetic Analysis of Complex Disease: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Genetic Analysis of Complex Disease»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
An up-to-date and complete treatment of the strategies, designs and analysis methods for studying complex genetic disease in human beings Genetic Analysis of Complex Diseases
Genetic Analysis of Complex Diseases
Genetic Analysis of Complex Diseases
Genetic Analysis of Complex Disease — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Genetic Analysis of Complex Disease», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
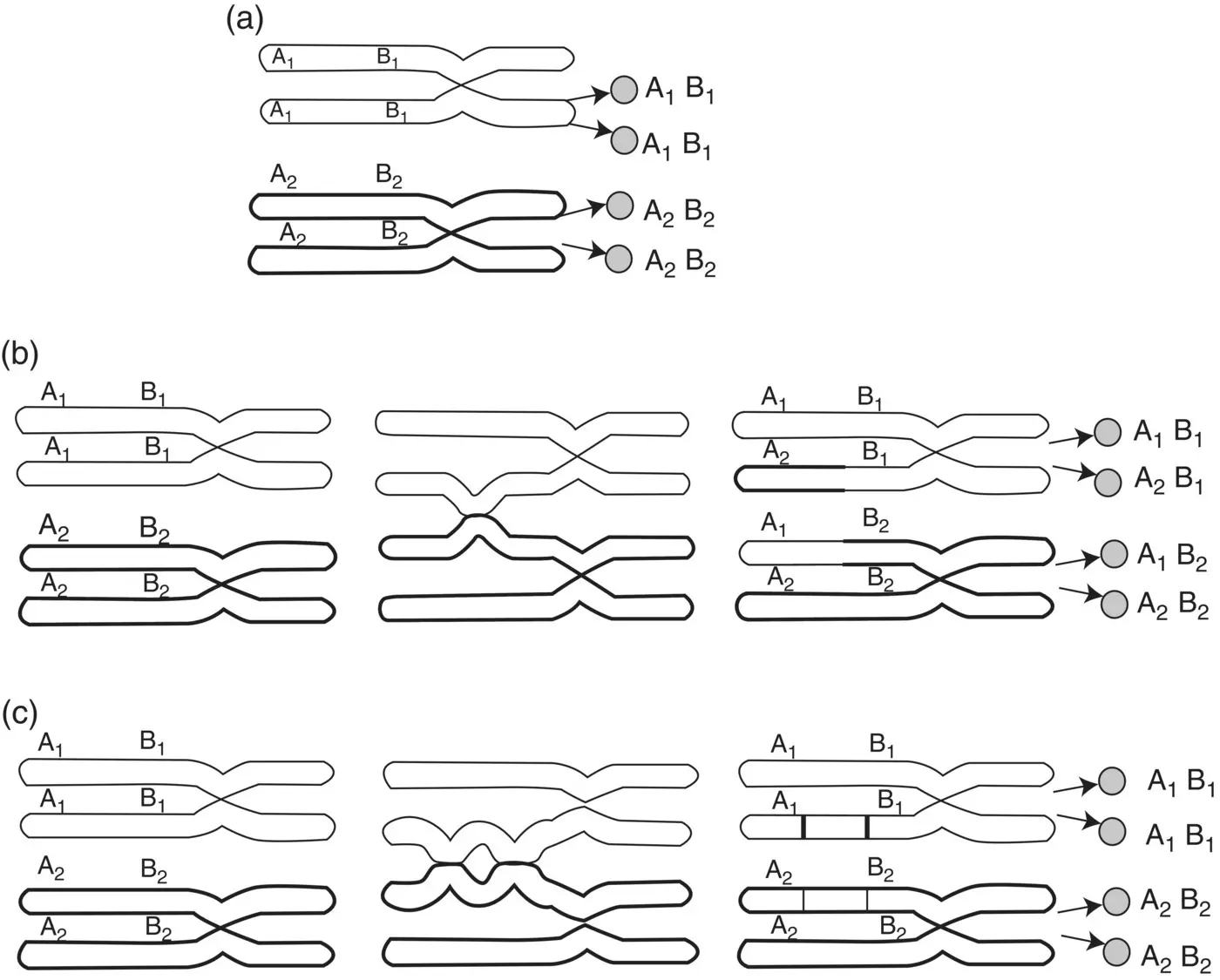
Figure 2.8 Genetic results of crossing over: (a) no crossover: A and B remain together after meiosis; (b) crossover between A and B results in a recombination (A and B are inherited together on a chromosome, and A and B are inherited together on another chromosome); (c) double crossover between A and B results in no recombination of alleles.
( Source: Reprinted by permission from Jorde et al. (1995).)
Following recombination, at least two of the four chromatids become unique, unlike those of the parent. The cellular division process that occurs ensures that one paternal homologue and one maternal homologue are transmitted to each of two diploid daughter cells. This cell division marks the end of meiosis I. The process of genetic recombination helps to preserve genetic variability within a species by allowing for virtually limitless combinations of alleles in the transmission from parent to offspring. Estimates of genetic recombination can also predict distance between two loci: The closer two loci are to one another, the less chance for recombination between them. The frequency of recombination is not uniform through the genome. Some areas of some chromosomes have increased rates of recombination (hot spots), while others have reduced rates of recombination (cold spots). For instance, recombination frequencies may vary between sexes or may vary depending on whether the loci are at the telomere or centromere of the chromosome.
The second phase of meiosis is identical to a mitotic (somatic cell) division, in which genetic material is transmitted equally, identically, and without recombination to daughter cells. However, in contrast to a mitotic division, which yields two identical diploid daughter cells, the end result of the entire meiotic process in sperm cells is four haploid daughter cells with chromosomal haplotypes different from those originally present in the parent; in egg cells, the final outcome is a single haploid daughter cell, with the remainder of the genetic material lost because of the formation of nonviable polar bodies.
The fundamental differences between meiosis and mitosis are summarized in Table 2.2.
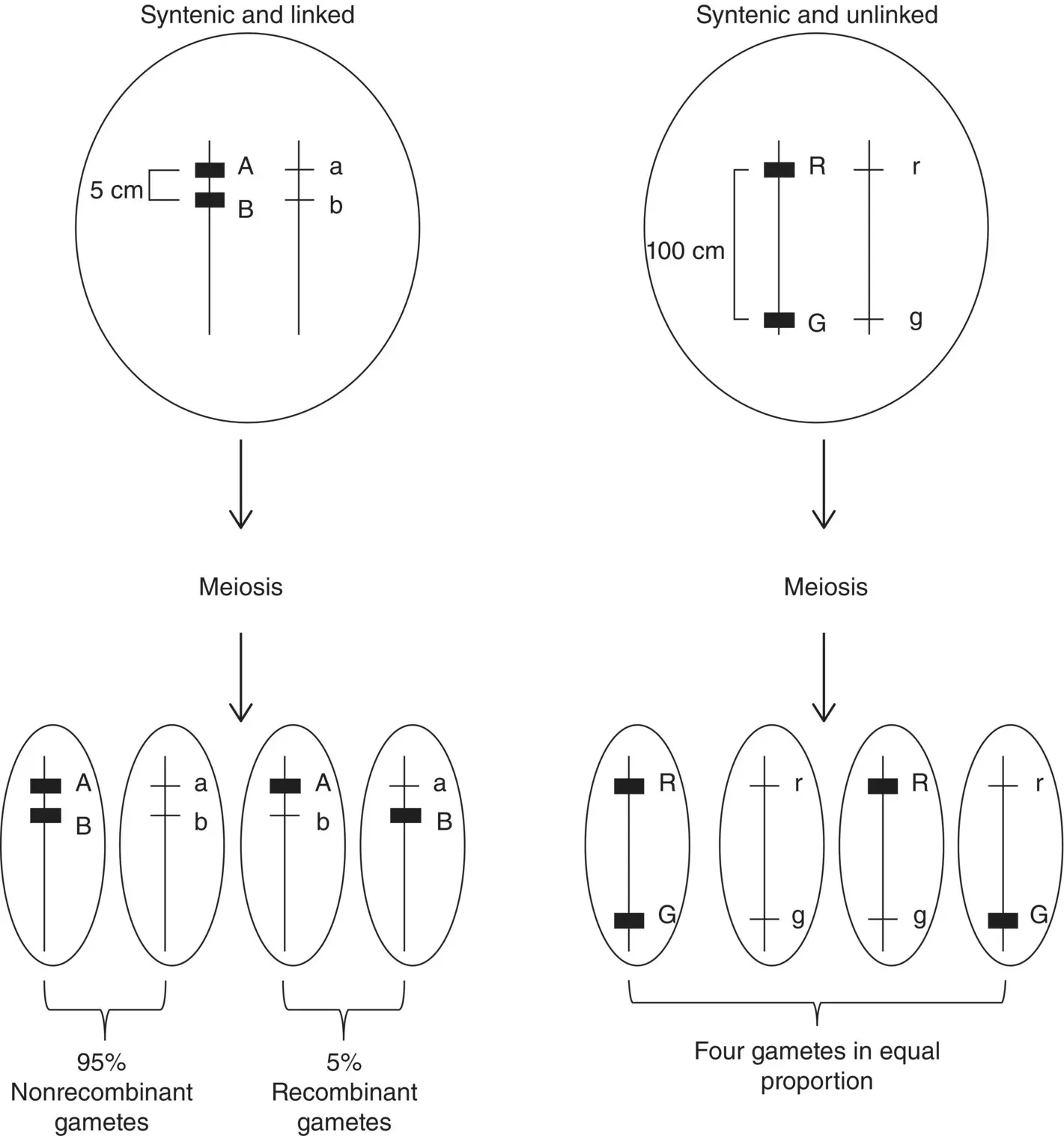
Figure 2.9 Genes that are on the same chromosome (syntenic) may be unlinked or linked. Loci A/a and B/b are syntenic and linked because their physical distance is short enough that recombination is less likely to occur between them, resulting in a lower recombinant to nonrecombinant gamete ratio. Loci R/r and G/g are syntenic and unlinked because their physical distance is great enough that a recombination event will occur between them during meiosis resulting in equal proportions of recombinant and nonrecombinant gametes.
Table 2.2 Differences between meiosis and mitosis.
| Meiosis | Mitosis | |
|---|---|---|
| Purpose | Produce gametes; ensure or produce genetic variability through recombination | Replace somatic cells |
| Location | Gonads | Body cells |
| Number of cell divisions per cycle | Two: meiosis I and meiosis II (latter identical to mitosis) | One |
| Chromosome number in daughter cells | Halved from parental complement to produce gametes; resultant cells are haploid | Identical to parental complement; resultant cells are diploid |
| Recombination | Occurs in diplotene of prophase I of meiosis I | Occurs rarely, usually result of abnormality |
When Genes and Chromosomes Segregate Abnormally
Failure of meiosis at either phase (meiosis I or II) is termed nondisjunction and leads to aneuploidy, or abnormal chromosomal complements. The most well‐known aneuploidy is Down syndrome. Most cases of Down syndrome are caused by an extra copy of chromosome 21, but ~5% of cases are caused by a translocation such that the trisomy is not of the whole chromosome. Down syndrome is often called trisomy 21 because most individuals with Down syndrome have a total of 47 chromosomes, with three copies of chromosome 21. A monosomy, or the absence of a second member of a chromosome pair, is rarely viable. A noted exception is Turner syndrome, in which a female has a total of 45 chromosomes, including a single X chromosome. Nondisjunction events that lead to abnormal chromosome complements may occur in the egg or the sperm. However, in contrast to male sperm, which are produced throughout the lifespan, female egg cells are suspended in meiosis II since shortly after that female’s conception. These eggs are subject to aging over the female lifespan, and errors of nondisjunction are more likely to occur.
Triploidy and tetraploidy are the terms for the presence of one or two entire extra sets of chromosomes, leading to a total of 69 or 92 chromosomes, respectively. These anomalies, which usually are not viable in humans, are due to errors in fertilization such as dispermy (two sperm fertilizing an ovum) or failure of the ovum’s polar body to separate.
Inheritance Patterns in Mendelian Disease
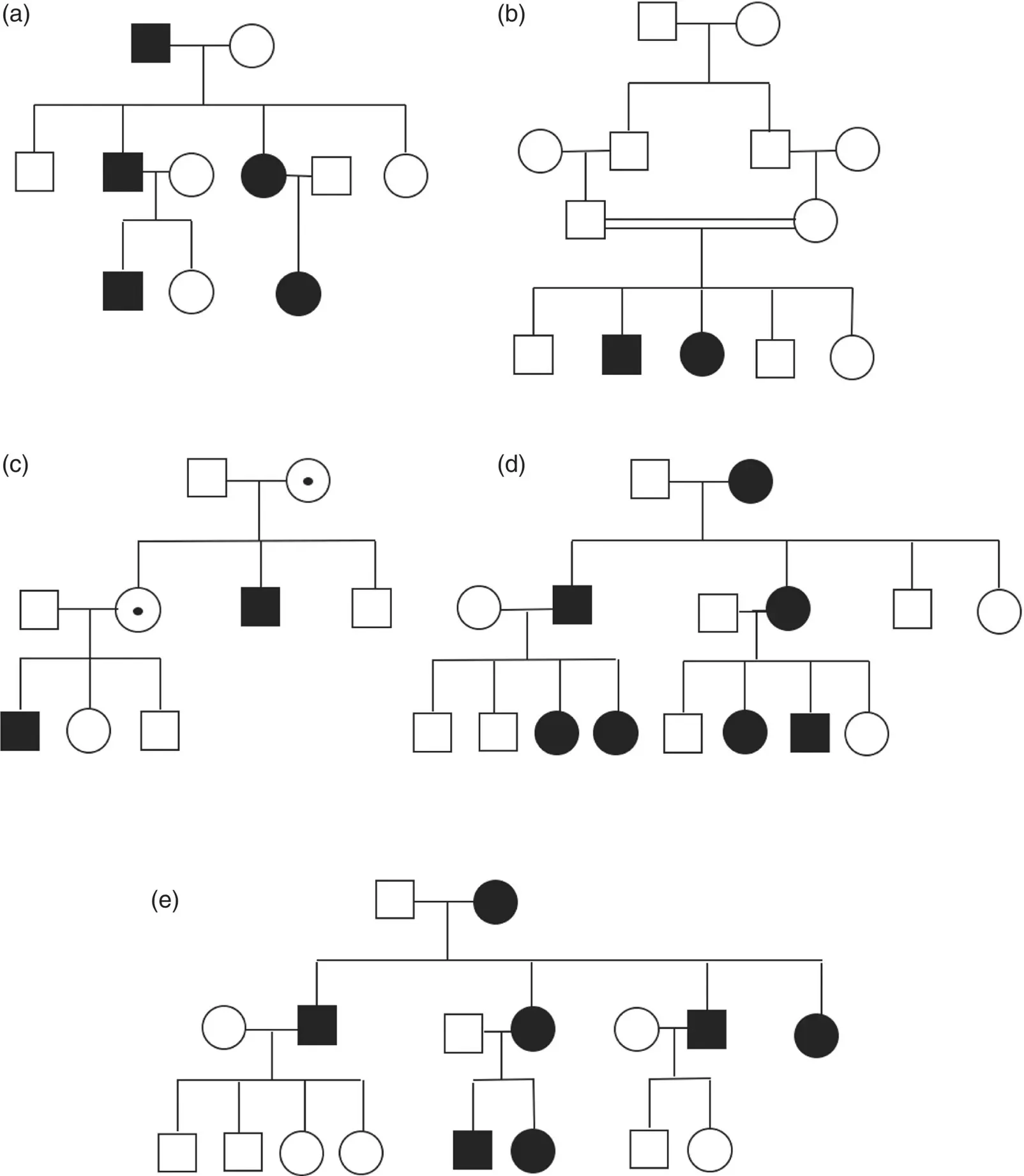
Many of the traits Mendel studied in his peas were inherited in a simple dominant or recessive fashion, a manner in which many human traits and disease are also inherited. Characteristics of each of the different Mendelian inheritance patterns are summarized in Table 2.3. Pedigrees representing these well‐known patterns of inheritance are shown in Figure 2.10.
Autosomal Recessive
A condition described as autosomal recessive maintains that the causative gene is located on an autosome and that both copies of the gene must harbor a causative allele. In most cases of recessive conditions, it is correctly assumed that each parent of an offspring with the trait carries a single causative allele. Carriers for recessive conditions do not typically show phenotypic features. Recessive conditions occur more commonly in offspring of consanguineous unions or in populations with high frequency of carriers for a particular condition.
Autosomal Dominant
In a dominant condition, a genetic mutation in a single copy of a gene is sufficient to cause a disease or trait. Dominant conditions may be inherited through an affected parent or alternatively may have occurred as the result of a de novo genetic mutation. Some de novo dominant mutations have been shown to be associated with increased paternal age (Friedman 1981). Some dominant conditions may exhibit reduced penetrance, in which a person carries a causative gene but does not exhibit any manifestations of the condition. On the other hand, variable expressivity refers to the range of features that may be observed in individuals with the same condition. Codominance, in which a trait is expressed from both alleles, has also been observed in certain traits, for example, in AB blood type.
Table 2.3 Hallmarks of Mendelian inheritance patterns of different types.
| Inheritance pattern | Examples | Transmission features | Recurrence risk | Prevalence in population | Other critical features |
|---|---|---|---|---|---|
| Autosomal dominant | Marfan syndrome; neurofibromatosis; myotonic dystrophy | Transmitted from affected parent to affected offspring (vertical transmission) male‐to‐male possible transmission; de novo mutations may occur | For each offspring of affected parent, risk to child to inherit disease gene is 50% | p 2+ 2pq | Reduced penetrance may be observed |
| Autosomal recessive | Sickle cell anemia; cystic fibrosis | Carrier parents generally unaffected | For carrier parents, risk for each subsequent child is 25% | q 2 | Consanguinity considered |
| X‐linked | Duchenne muscular dystrophy; fragile X syndrome; hemophilia | No male‐to‐male transmission; de novo mutations may rarely occur | 50% of offspring of carrier female have trait (if male, affected, if female carrier); all female offspring of affected male are carriers | Females: q 2; males: q | Females may show sub‐clinical, atypical, or fully penetrant features of the condition. Non‐random X inactivation may contribute to more severe female phenotype. |
| Y linked | Genes SRY and TDF, important in sex determination, are on the Y chromosome; no known diseases are located on Y | Exclusively male‐to‐male transmission | All sons of affected males are affected; no daughters of affected males are affected | Females: 0; males: q | Male‐determining genes are located just proximal to pseudoautosomal region on Y chromosome; faulty recombination in pseudoautosomal region can lead to errors in sex determination |
| Autosomal codominant | MN blood group; microsatellite repeat markers | Each allele confers measurable component to phenotype | Varies according to mating type | Genotypes expected to occur in Hardy–Weinberg proportions of p 2, 2pq, and q 2 | |
| Mitochondrial | Leber’s optic atrophy; KSS (Kearns–Sayre syndrome); MELAS (mitochondrial myopathy, encephalopathy, lactic acidosis and stroke‐like episodes); | Exclusively maternal transmission through maternal mitochondria | All offspring of affected females are at risk to inherit mutation (may be affected or carrier). Proportion of affected offspring is variable based on maternal heteroplasmy. Offspring of affected male not at risk to inherit mutation. | Heteroplasmy may determine phenotypic severity. Majority of mitochondrial diseases are due to mutations in the nuclear genome rather than the mitochondrial genome and follow autosomal recessive inheritance pattern. |
Интервал:
Закладка:
Похожие книги на «Genetic Analysis of Complex Disease»
Представляем Вашему вниманию похожие книги на «Genetic Analysis of Complex Disease» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.











Обсуждение, отзывы о книге «Genetic Analysis of Complex Disease» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.