Joseph J. Torres - Life in the Open Ocean
Здесь есть возможность читать онлайн «Joseph J. Torres - Life in the Open Ocean» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Life in the Open Ocean
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Life in the Open Ocean: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Life in the Open Ocean»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Life in the Open Ocean: The Biology of Pelagic Species
Life in the Open Ocean: The Biology of Pelagic Species
Life in the Open Ocean — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Life in the Open Ocean», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
(2.1) 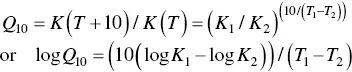
where K 1and K 2are velocity constants corresponding to temperatures T 1and T 2. Reaction velocity is generally used instead.
For virtually all rate functions in which we are interested, the biological rate increases by a factor of approximately 2 for each 10 °C rise in temperature: that is, Q 10≈ 2. However, the Q 10of an animal’s metabolic rate varies slightly over a range of temperatures, being higher in the lower ranges. Therefore, when providing Q 10for metabolic rates, it is imperative to also provide the temperature range over which the measurements were taken.
The fact that metabolic rate doubles with a temperature change of 10 °C (or halves with a 10 °C drop) has a profound effect on ectothermic species. A vertical migrator in the tropical ocean swimming from a depth of 500 m to near surface waters at sunset (a common occurrence, as we shall see) will encounter changes of >10 °C on its way up and again on its way back to depth. As a consequence, it will endure profound changes in its metabolic rate during each leg of its excursion.
In 1914, August Krogh, the father of comparative physiology, first attempted to define a pattern for the change in Q 10with temperature by subjecting a narcotized goldfish to temperatures ranging between 0 and 25 °C and measuring its oxygen consumption rate. The curve he derived is called the “Normal Curve.” It was popularized considerably in later years when it was found that a similar relationship between metabolism and temperature existed for many species, with the exception of the large Q 10in the 0–5 °C temperature interval (e.g. Winberg 1956). Even Krogh stated that his Q 10value of 10.9 between 0 and 5 was “obviously erroneous.” The general trend was remarkably accurate though, as were the numbers generated above 5 °C. The Q 10’s this curve represents is shown below.
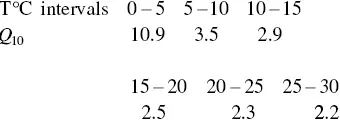
The Q 10approximation is a fundamental molecular response to temperature: it applies to chemical reactions taking place in a beaker as well as to rate processes in ectothermic species. However, it is not intuitively obvious why reaction rates should double for every increase in a temperature of 10 °C. The answer is in a concept termed “activation energy,” which was pioneered by the Swedish physical chemist Svante Arrhenius, and which earned him a Nobel Prize in 1903.
In the realm of physical chemistry, temperatures are expressed in the absolute temperature scale, in degrees Kelvin. A Kelvin degree is equal to a degree Celsius, but the scale begins at absolute zero, the temperature where all molecular motion ceases: −273 °C. Thus, a temperature of 0 °C is equal to 273 K, and 20 °C is equal to 293 K; by convention, the degree symbol is not used for degrees Kelvin. The temperature range most relevant to the pelagic fauna, −2 to 40 °C (271–313 K), only covers about a 10% change of temperature on the absolute scale. In our range of concern, a change of 10 °C is roughly 3% of the absolute temperature. Why, then, do reaction rates double?
The breakthrough of Arrhenius was his idea that within a population of molecules, only a fraction have sufficient energy to be reactive: those that exceed the activation energy threshold ( Figure 2.3). The average thermal energy of the molecules gives us the temperature, but it is not the average that is most important, it is the proportion of molecules that have enough energy to exceed the activation energy threshold and be competent to react. When heat is added to a system, the proportion of molecules that exceeds the activation energy increases more quickly than the average temperature. In fact, an increase in temperature of 10 K results in a doubling of the fraction of molecules exceeding the activation energy. The activation energy concept thus explains the Q 10s we observe with biological rates.
Experimentation in the 1940s, 1950s, and 1960s further defined temperature responses as a function of time and acclimation period. Three general time courses were identified.
1 Direct responses of rate functions to changes in temperature persisting for hours: acute measurements
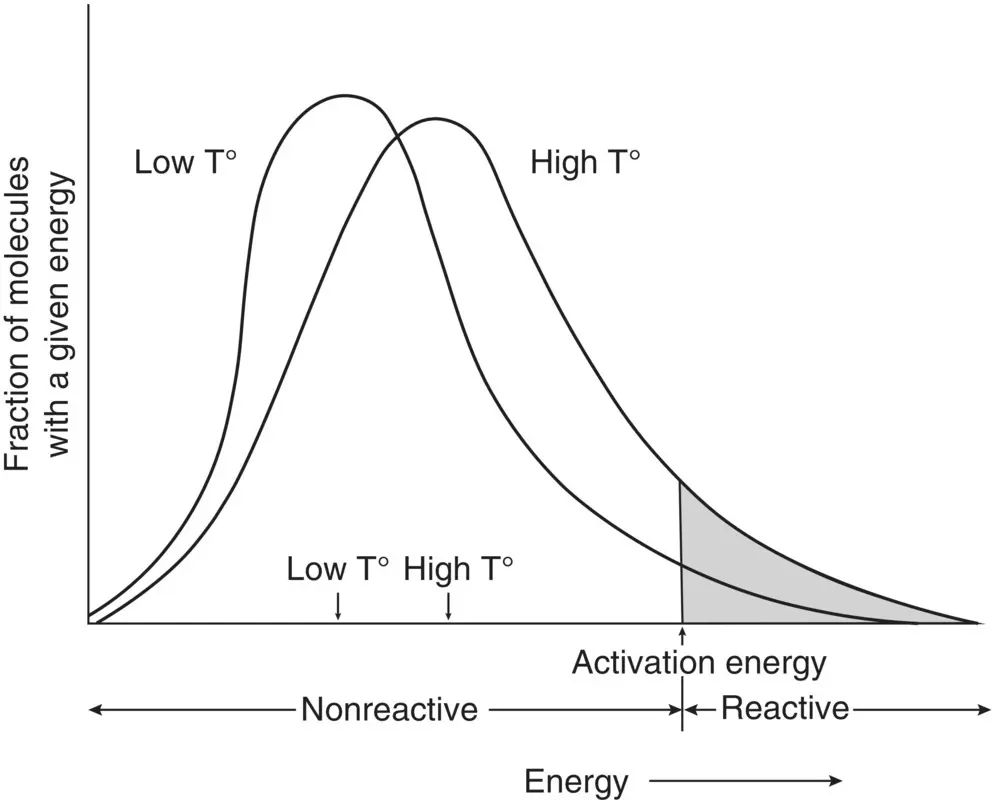
Figure 2.3 Energy distribution curves for a population of molecules at two different temperatures. Only those molecules having energies equal to or greater than the activation energy are reactive.
Source: Hochachka and Somero (2002), figure 7.1 (p. 296). Reproduced with the permission of Oxford University Press.
Rates measured in this way, with no acclimation period, reflect the short‐term flexibility of biological systems. In some cases, acutely measured responses to temperature can show Q 10values greatly different from 2. When metabolism is the rate being measured, such a response is termed “metabolic overshoot.” It would correspond to a “type 5” response described below. It is the result of a system that is still in the process of adjusting to a new temperature and it can happen in a transition to a warmer or colder temperature.
1 Compensatory acclimation to days or weeks of exposure: the acclimated response
An animal is acclimated only when its rate processes have stabilized to the new temperature. Acclimated animals were utilized in constructing the temperature tolerance polygon shown in Figure 2.2a.
1 Evolutionary adaptation through natural selection: climatic adaptation
Patterns of Thermal Acclimation
Animals moved to a new temperature and kept there for several days often show some compensation in their rate functions, especially metabolic rate. One of the classic treatments of how individuals adjust to a new temperature is that of Precht (1958) who identified five responses ( Figure 2.4). Animals were maintained initially at temperature T 2and moved to T 1or T 3. Metabolic rates were monitored during the acclimation process.
Type 4 acclimation: no change in rate occurs after time for acclimation (Q10 = 2–3).
Type 2 acclimation: the animal’s rate falls or rises to original rate (Q10 = 1)
Type 3 acclimation: somewhere in between (Q10 = 1–2)
Type 1 acclimation: overcompensation – rate lower at higher temperature (Q10 < 0)
Type 5 acclimation: reverse compensation (Q10 > 3)
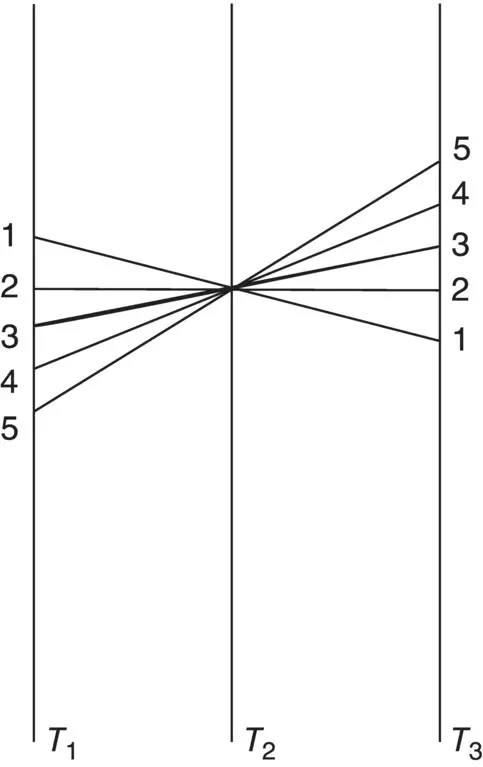
Figure 2.4 Precht’s patterns of temperature acclimation. Animals are acclimated at temperature T 2then transferred to a lower temperature ( T 1) or higher temperature ( T 3). The five acclimation types are described in the text.
Source: Prosser (1973), figure 9‐13 (p. 375). Reproduced with the permission of Saunders Publishing.
In reality, all five curves are rarely seen. In most cases, the response is type 3 or 4. However, type 2 has been seen in rare cases; it has been documented in some intertidal animals (Newell and Northcroft 1967). The Precht patterns are the simplest way of classifying trends in metabolism vs. temperature, but other useful schemes exist. Prosser (1973) describes species’ response to temperature using a family of metabolism vs. temperature (M‐T) curves instead of Precht’s three points. The advantage of Prosser’s scheme is that it can differentiate varied responses to lowered and raised temperature, and the reader is encouraged to examine Prosser’s curves.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Life in the Open Ocean»
Представляем Вашему вниманию похожие книги на «Life in the Open Ocean» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Life in the Open Ocean» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.