Stephen J. Bourke - Respiratory Medicine
Здесь есть возможность читать онлайн «Stephen J. Bourke - Respiratory Medicine» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Respiratory Medicine
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Respiratory Medicine: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Respiratory Medicine»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
, a team of distinguished physicians delivers a comprehensive and accessible overview of the essentials of respiratory medicine, including a review of respiratory anatomy and physiology, as well as the aetiology, epidemiology, symptoms, and management of a wide range of respiratory diseases.
This edition offers self-assessment exercises in each chapter and a range of clinical images and scans showing the critical features of each disease. The book also offers:
A thorough introduction to history taking, examination, and investigations Comprehensive explorations of respiratory diseases, including upper respiratory tract infections and influenza, pneumonia, and tuberculosis Practical discussions of bronchiectasis, lung abscess, cystic fibrosis, asthma, and chronic obstructive pulmonary disease In-depth examinations of lung transplantation A companion website featuring figures, key points, and interactive self-assessment questions Perfect for medical students and respiratory nurses,
will also earn a place in the libraries of early-career medical doctors and residents with an interest in respiratory medicine.
Respiratory Medicine — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Respiratory Medicine», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
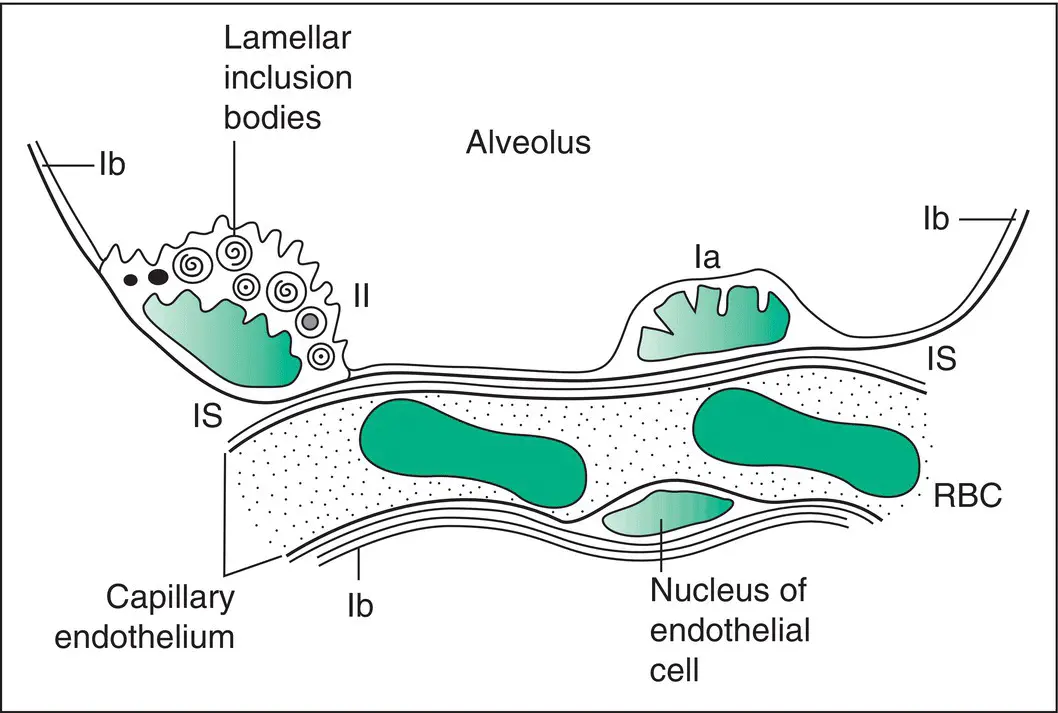
Figure 1.3 Structure of the alveolar wall as revealed by electron microscopy. la, type I pneumocyte; lb, flattened extension of type I pneumocyte covering most of the internal surface of the alveolus; II, type II pneumocyte with lamellar inclusion bodies, which are probably the site of surfactant formation; IS, interstitial space; RBC, red blood corpuscle. Pneumocytes and endothelial cells rest upon thin continuous basement membranes, which are not shown.
Physiology
The core business of the lungs is to bring oxygen into the body and to take carbon dioxide out. The deceptively simple act of ‘breathing’ comprises two quite distinct processes.
1 Ventilation. The movement of air in and out of the lungs (between the outside world and the alveoli).
2 Gas exchange. The exchange of oxygen and carbon dioxide between the airspace of the alveoli and the blood.
Ventilation continues throughout life, largely unconsciously, coordinated by a centre in the brain stem. The factors that regulate the process, ‘the control of breathing’, will also be considered here. Gas exchange happens automatically (by diffusion) if blood and inspired air are brought into close proximity.
Ventilation
To understand this process, we need to consider the muscles that ‘drive the pump’ and the resistive forces they have to overcome. These forces include the inherent elastic property of the lungs and the resistance to airflow through the bronchi (airway resistance).
The muscles that drive the pump
Inspiration requires muscular work. The diaphragm is the principal muscle of inspiration. At the end of an expiration, the diaphragm sits in a high, domed position in the thorax ( Fig. 1.4). To inspire, the strong muscular sheet contracts, stiffens and tends to push the abdominal contents down. There is variable resistance to this downward pressure by the abdomen, which means that in order to accommodate the new shape of the diaphragm, the lower ribs (to which it is attached) also move upwards and outwards. (When airway resistance is present, as in asthma or chronic obstructive pulmonary disease [COPD], the situation is very different; see Chapters 2& 11.) The degree of resistance the abdomen presents can be voluntarily increased by contracting the abdominal muscles; inspiration then leads to a visible expansion of the thorax, rather than a distension of the abdomen (try it). The resistance may also be increased by abdominal obesity. In such circumstances, there is an involuntary limitation to the downward excursion of the diaphragm and, as the potential for upward movement of the ribs is limited, the capacity for full inspiration is diminished. This inability to fully inflate the lungs is an example of a restrictive ventilatory defect(see Chapter 3).
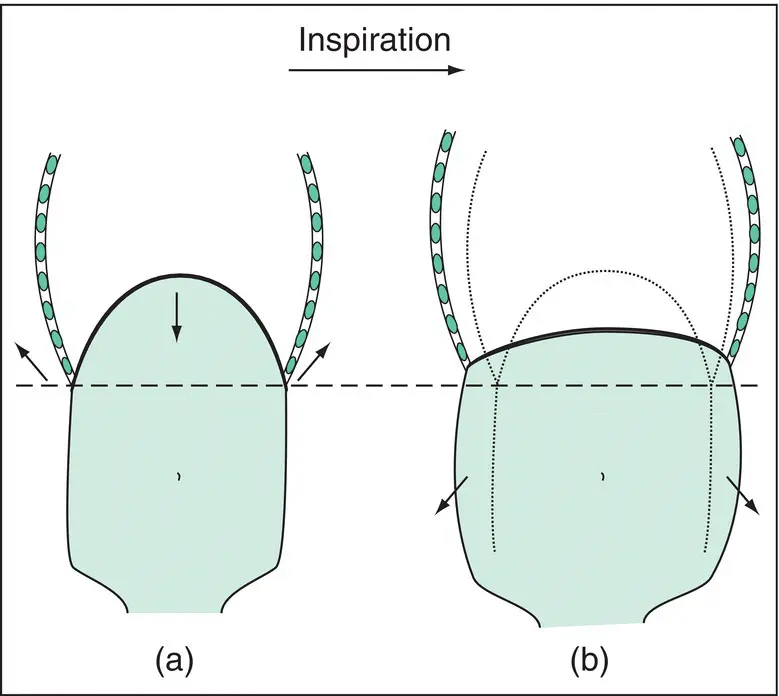
Figure 1.4 Effect of diaphragmatic contraction. Diagram of the ribcage, abdominal cavity and diaphragm showing the position at the end of resting expiration (a). As the diaphragm contracts, it pushes the abdominal contents down (the abdominal wall moves outwards) and reduces pressure within the thorax, which ‘sucks’ air in through the mouth (inspiration). (b) As the diaphragm shortens and descends, it also stiffens. The diaphragm meets a variable degree of resistance to downward discursion, which forces the lower ribs to move up and outward to accommodate its new position.
Other muscles are also involved in inspiration. The scalene muscles elevate the upper ribs and sternum. These were once considered, along with the sternocleidomastoids, to be ‘ accessory muscles of respiration’, only brought into play during the exaggerated ventilatory effort of acute respiratory distress. Electromyographic studies, however, have demonstrated that these muscles are active even in quiet breathing, although less obviously so.
The intercostal muscles bind the ribs to ensure the integrity of the chest wall. They therefore transfer the effects of actions on the upper or lower ribs to the whole ribcage. They also brace the chest wall, resisting the bulging or in‐drawing effect of changes in pleural pressure during breathing. This bracing effect can be overcome to some extent by the exaggerated pressure changes seen during periods of more extreme respiratory effort, and in slim individuals intercostal recessionmay be observed as a sign of respiratory distress.
Whilst inspiration is the result of active muscular effort, quiet expiration is a more passive process. The inspiratory muscles steadily release their contraction and the elastic recoil of the lungs brings the tidal breathing cycle back to its start point. Forced expiration, however – either volitional or as in coughing – requires muscular effort. The abdominal musculature is the principal agent in this.
The inherent elastic property of the lungs
Lung tissue has a natural elasticity. Left to its own devices, a lung would tend to shrink to little more than the size of a tennis ball. This can sometimes be observed radiographically in the context of a complete pneumothorax(see Chapter 16). The lung’s tendency to contract is counteracted by the semi‐rigid chest wall, which itself has a tendency to spring outward from its usual position. At the end of a normal tidal expiration, the two opposing forces are nicely balanced and no muscular effort is required to hold this ‘neutral’ position. Breathing at close to this lung volume (normal tidal breathing) is therefore relatively efficient and minimises work. It is rather like gently stretching and relaxing a spring from its neutral, tension‐free position. In some diseases (asthma or COPD), tidal ventilation is obliged to occur at higher lung volumes (see Chapter 3). Breathing then is rather like stretching and relaxing a spring that is already under a considerable degree of tension. The work of breathingis therefore increased, a factor that contributes to the sensation of breathlessness. Test this yourself: take a good breath in and try to breathe normally at this high lung volume for a minute.
The natural tendencies for the chest wall to spring outwards and the lung to contract down present opposing forces, which generate a negative pressure within the pleural space. This negative pressure (‘vacuum’) maintains the lung in its stretched state. Clearly, at higher lung volumes, the lung is at greater stretch and a more negative pleural pressure is required to hold it in position. The relationship between pleural pressure (the force on the lung) and lung volume can be plotted graphically ( Fig. 1.5). The lung does not behave as a perfect spring, however. You may recall that the length of a spring is proportional to the force applied to it (Hooke’s law). In the case of the lung, as its volume increases, greater and greater force is needed to achieve the same additional increase in volume; that is, the lung becomes less ‘compliant’ as its volume increases. Lung complianceis defined as ‘the change in lung volume brought about by a unit change in transpulmonary (intrapleural) pressure’.
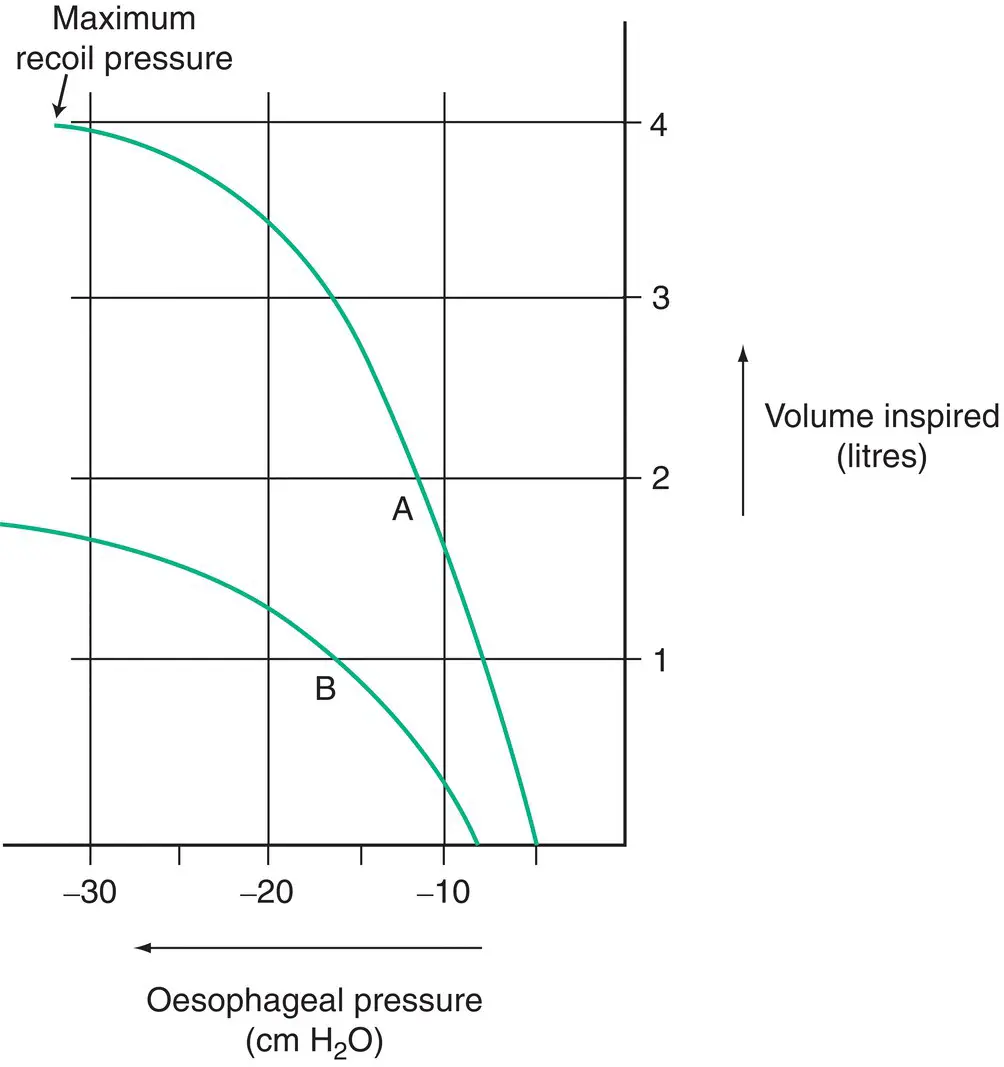
Figure 1.5 Graph of (static) lung volume against oesophageal pressure (a surrogate for intrapleural pressure). In both subjects A and B, we see that lung compliance – the change in lung volume per unit change in intrapleural pressure (or slope of the curve) – is reduced at higher lung volumes. A: normal individual. B: individual with reduced lung compliance, such as lung fibrosis.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Respiratory Medicine»
Представляем Вашему вниманию похожие книги на «Respiratory Medicine» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.











Обсуждение, отзывы о книге «Respiratory Medicine» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.