Kirk N. Gelatt - Essentials of Veterinary Ophthalmology
Здесь есть возможность читать онлайн «Kirk N. Gelatt - Essentials of Veterinary Ophthalmology» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.
- Название:Essentials of Veterinary Ophthalmology
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Essentials of Veterinary Ophthalmology: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Essentials of Veterinary Ophthalmology»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Ophthalmic foundations: ophthalmic development and structure, physiology of the eye and vision, and ocular pharmacology and therapeutics Canine ophthalmology: canine orbit (disease and surgery), canine eyelids (disease and surgery), canine lacrimal apparatus (tear secretion and drainage), canine cornea (diseases and surgery) and canine glaucoma Other species: feline ophthalmology, equine ophthalmology, and food and fiber animal ophthalmology Ophthalmic and systemic diseases: comparative neuro-ophthalmology and systemic disease and the eye
is a useful guide for veterinary students and practitioners looking to build out their core foundations of knowledge within their specific programs of study and disciplines.
Essentials of Veterinary Ophthalmology — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Essentials of Veterinary Ophthalmology», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Cholinergic regulation of AH formation and composition is similarly ambiguous. For example, parasympathomimetic nerve stimulation or drugs have been demonstrated to increase, decrease, or not change the rate of AH production; these differences are likely due to species and technique‐related effects. Cholinergic agents may regulate amino acid transport from the blood to the AH as well as modulate inorganic ion concentrations within the AH. In aggregate, these studies suggest that the influence of parasympathetic drugs such as pilocarpine is relatively minor in AH formation and that their efficacy in decreasing IOP is likely due to increased AH outflow.
Aqueous Humor Outflow
AH dynamics involve the balance between production (i.e., active secretion) and outflow via the conventional and nonconventional routes ( Figure 2.7). Conventional outflow consists of AH flow through the corneoscleral TM, while the nonconventional route involves uveoscleral outflow. Depending on the species, ~50–95% of AH drains through the TM via conventional outflow ( Table 2.6).
Structural and Biomechanical Attributes
The TM is a complex, three‐dimensional structure comprising cells supported by an intricate ECM. The TM can be divided into three portions: uveal, the innermost portion; corneoscleral, the middle region; and the juxtacanalicular zone, the outermost section nearest the sclera (probably the site of greatest outflow resistance). The pore size of each TM zone decreases progressively from inward to outward, resulting in progressive increases in outflow resistance to the passage of AH. The juxtacanalicular zone consists of several endothelial cell layers that produce a matrix comprising GAGs, collagen, fibronectin, and other glycoproteins in which these cells are embedded. Thus, the juxtacanalicular zone, which is immediately adjacent to Schlemm's canal in primates or the angular aqueous plexus (AAP) in most domestic animals, is thought to be the major site of aqueous outflow resistance.
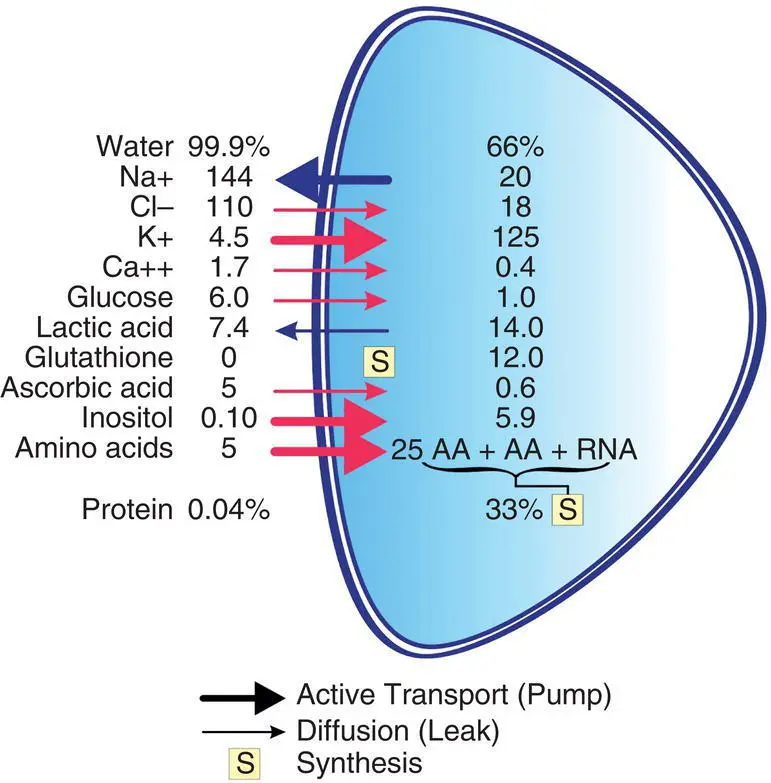
Figure 2.7 Chemical composition of the aqueous humor and lens. Water and protein are expressed as percentages of lens weight. Na +, Cl −, K +, and Ca 2+ions are expressed in microequivalent per milliliter of lens water. Other compounds are expressed in micromole per gram of lens weight or micromole per milliliter of aqueous humor. AA, amino acid; RNA, ribonucleic acid.
Table 2.6 Estimates of aqueous humor dynamics in selected species.
| Dog | Cat | Rabbit | Cow | Horse | Nonhuman primate | |
|---|---|---|---|---|---|---|
| Estimated normal IOP (mmHg) | 15–18 | 17–19 | 15–20 | 20–30 | 17–28 | 13–15 |
| “ C ” outflow (μl/mmHg/min by tonography) | 0.24–0.30 | 0.27–0.32 | 0.22–0.28 | — | 0.90 | 0.24–0.28 |
| Uveoscleral outflow (μl/min) | 15% | 3% | 13% | — | — | 30–65% |
| Episcleral venous pressure (mmHg) | 10–12 | 8 | 9 | — | — | 10–11 |
| Aqueous formation (μl/min) | 5.22 | 6.00–7.00 | 1.84 | — | — | 2.75 |
Other studies suggest that the main site of resistance to outflow is the endothelial lining of the AAP and its ECM. However, the site of filtration may be different from the site of flow resistance. AH transport through the endothelium of the AAP (or Schlemm's canal in nonhuman primates and domestic chickens) is thought to occur via transcellular pores, large vacuoles, or pinocytotic vesicles. However, paracellular routes between the endothelial cells of Schlemm's canal have also been proposed and may be pressure sensitive, particularly at higher IOPs.
Fluid Dynamics
As the ciliary body produces AH, the tissues comprising the ICA resist AH outflow, thus generating IOP. Steady‐state IOP occurs when the rates of AH inflow and outflow are equivalent. The AH exits the eye by passive bulk flow via two routes in the ICA:
1 The traditional or conventional pathway, which involves passage through the TM, AAP, scleral venous plexus, veins of the episclera and conjunctiva (anterior) or vortex veins (posterior), and systemic venous circulation.
2 The uveoscleral or nonconventional pathway, which involves passage through the iris root, anterior face of the ciliary body muscle, supraciliary or suprachoroidal space, and out through the sclera (and perhaps the optic nerve head).
The traditional pathway is dependent on IOP, while the uveoscleral pathway is not as long, since IOP is greater than 7–10 mmHg. At very low IOP, the net pressure gradient across the nonconventional pathway declines, so that uveoscleral outflow subsequently decreases. It is unknown why uveoscleral outflow is largely independent of IOP, but it may relate to complex relationships between pressure and resistance between the fluid compartments and the soft tissues that comprise this route. For example, the pressure gradient between the anterior chamber and suprachoroid is independent of IOP; thus, fluid flow between these compartments is also IOP independent. Uveoscleral outflow is primarily impacted by the state of the ciliary body and by the hydrostatic pressure difference between the anterior chamber and the suprachoroidal space. Contraction of the ciliary body musculature decreases unconventional outflow, possibly by reducing the extracellular spaces; in turn, relaxation increases outflow via this route. Thus, pilocarpine, a parasympathomimetic drug, and atropine, a parasympatholytic drug, will decrease and increase uveoscleral outflow by contracting and relaxing the ciliary body musculature, respectively. Because of general venous pressure, IOP in the eye under general anesthesia will decrease to only 10–12 mmHg.
Table 2.7 Aqueous humor dynamics formulas.
| I | F in= F at+ F uf |
| F = flow (μl/min) | |
| F in= total AH inflow | |
| F at= inflow from active transport | |
| F uf= inflow from ultrafiltration | |
| II | F out= F trab+ F uveo |
| F out= total AH outflow | |
| F trab= outflow via the TM | |
| F uveo= outflow via the uveoscleral pathway | |
| III | C total= C trab+ C uv+ C pseudo |
| C = facility or conductance of flow (μl/min/mmHg) | |
| C total= total AH outflow facility | |
| C trab= facility of outflow via the TM | |
| C uv= facility of outflow via the uveoscleral pathway | |
| C pseudo= pseudofacility | |
| III | At steady state, F = F in= F out |
| IV | F = C trab( P i− P e) (Goldmann equation) |
| P = pressure (mmHg) | |
| P i= IOP | |
| P e= episcleral venous pressure | |
| V | F in= C trab( P i− P e) + F uveo |
| VI | P i= P e+ ( F in− F uveo)/ C trab |
Formulas can be used to describe the formation and drainage of AH ( Table 2.7). Episcleral venous pressure or the “backpressure” created by the venous portion of the conventional pathway in the AAP or Schlemm's canal constitutes approximately 50–75% of the resistance (10–12 mmHg) that determines IOP. While minor anatomical variations in the venous system exist between species, results of pressure studies in humans, nonhuman primates, rabbits, and dogs reveal episcleral venous pressure to be between 8 and 12 mmHg. Arteriovenous anastomoses within the episcleral vasculature have been demonstrated in the rabbit, dog, owl monkey, and cynomolgus monkey. These vascular shunts may function in rabbits and dogs, where the episcleral vasculature appears to lack a capillary system, and in the monkey species as an emergency system to elevate IOP after globe perforation or to retrogradely flush the outflow channel. Episcleral venous pressure can be measured by direct cannulation (using very fine glass pipettes) or indirect partial to complete compression schemes (using a string‐gauge system or a fluid‐filled chamber). Results of limited studies indicate that the volume of the anterior chamber directly relates to the rate of aqueous outflow, so that animals with large eyes have faster outflow rates per minute. The resistance to aqueous outflow may be inversely proportional to the facility of outflow ( C total).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Essentials of Veterinary Ophthalmology»
Представляем Вашему вниманию похожие книги на «Essentials of Veterinary Ophthalmology» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.











Обсуждение, отзывы о книге «Essentials of Veterinary Ophthalmology» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.