Иван Павлов - Лекции о работе больших полушарий головного мозга
Здесь есть возможность читать онлайн «Иван Павлов - Лекции о работе больших полушарий головного мозга» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 2016, ISBN: 2016, Издательство: Литагент 5 редакция, Жанр: psy_generic, Биология, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Лекции о работе больших полушарий головного мозга
- Автор:
- Издательство:Литагент 5 редакция
- Жанр:
- Год:2016
- Город:Москва
- ISBN:978-5-699-81061-1
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Лекции о работе больших полушарий головного мозга: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Лекции о работе больших полушарий головного мозга»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
В своих знаменитых «Лекциях о работе больших полушарий головного мозга» он описывает основные нервные процессы, разъясняет понятие рефлекса, проливает свет на такие явления, как сновидения, гипноз и нарушения сна. Научное наследие Ивана Павлова определило облик современной физиологии и ряда смежных отраслей – биологии и медицины, психологии и педагогики.
Лекции о работе больших полушарий головного мозга — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Лекции о работе больших полушарий головного мозга», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
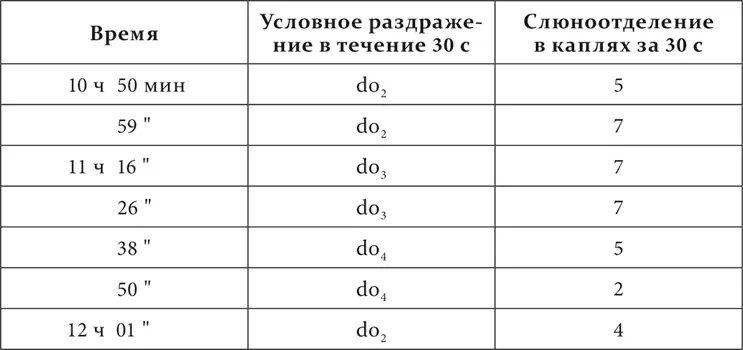
Та же собака спустя 3 дня.


Таким образом, дальнейшее основание для функциональной мозаичности коры надо видеть во взаимной индукции нервных процессов. Затем в этих же опытах мы занимались определением районов (полей) как раздражительного, так и тормозного процессов, а также возможных индифферентных, промежуточных между деятельными районами.
«Мы знаем из факта первоначальной генерализации условных раздражителей, что раздражение сперва непременно иррадиирует из исходного пункта, захватывает клетки и других рецепторных элементов»
На собаках с тоновой мозаикой мы пробовали другие тоны, находящиеся между нашими положительными и отрицательными раздражителями, и наблюдали их эффекты. Положительные эффекты в виде наступающего слюноотделения, конечно, были очевидны прямо. Что касается до нулевых, то надо было разобраться особыми вариациями опыта в их индифферентном или тормозном значении. Мы испытали их ради этого или на их последовательное торможение, или на их положительную индукцию.
Опыт 9.X.1925

Опыт 14.X.1925
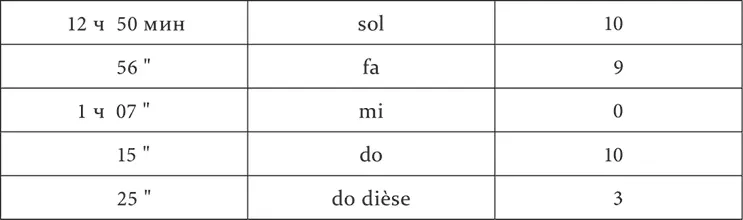
Опыт 20.X.1925
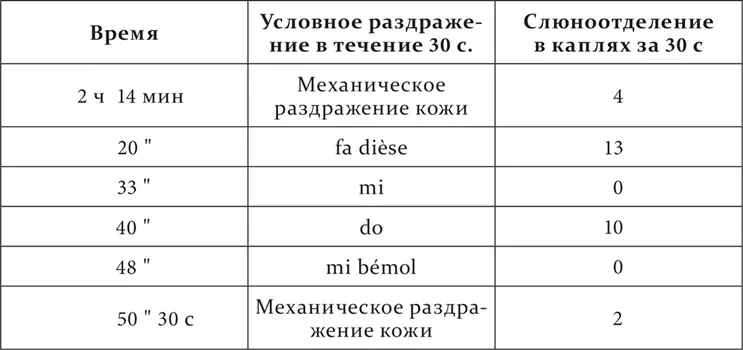
Мы видим, что из расположенных между положительным do и отрицательным mi промежуточные тон re и полутоны do dièse и mi bémol оказались разного значения. Полутон do dièse имел, хотя и уменьшенный, но положительный эффект, т. е. принадлежал к району положительного do. Тон re и полутон mi bémol, по-видимому, были одинаковы по действию, дав нуль. Но испытание их на последовательное торможение обнаружило их разницу. 2 минуты спустя после применения полутона mi bémol условное механическое раздражение потеряло в своем обыкновенном эффекте 50 %, т. е. было заторможено, после же тона re в тех же условиях времени осталось без изменения. Следовательно, полутон mi bémol бесспорно находился в районе отрицательного mi, тон же re или был совершенно индифферентным, или заторможен в гораздо меньшей степени. Последнее надо было решить более тонкими опытами, и потому вопрос об истинно индифферентных пунктах подвергается дальнейшему исследованию.
В этих же опытах исследовалось также взаимодействие пунктов при непосредственном переходе с раздражением от одних положительных пунктов к другим положительным, от одних отрицательных к другим отрицательным, от положительных к отрицательным, и наоборот. Но результаты большей частью получались такой сложности, что мы были вынуждены решать ту же задачу в более упрощенной обстановке.
Как указано было еще в первой лекции и упомянуто опять в начале этой, функциональная мозаика коры не только постоянно может пополняться, но и подлежит частой переделке, т. е. одни и те же пункты коры связываются то с одной, то с другой физиологической деятельностью организма. К последнему пункту относится у нас только одно исследование (опыты С. С. Фридемана ).
Одни и те же агенты делались сначала условными пищевыми раздражителями, а затем переделывались в кислотные, и наоборот – прежние кислотные раздражители превращались в пищевые. У двух собак первоначальный условный пищевой раздражитель был превращен в условный кислотный, а у третьей – кислотный в пищевой. Производилось это, конечно, так, что условный раздражитель сопровождался теперь другим безусловным раздражителем. Общий ход дела представляется в следующем виде. Условный раздражитель быстро, даже в течение одного опыта, теряет свой прежний слюноотделительный эффект и становится нулевым на более или менее значительный срок и затем снова приобретает слюногонное действие – и это уже был другой рефлекс, как показывало резкое изменение в составе слюны подчелюстной железы. Вся эта процедура требует около 30 сочетаний условного раздражителя с новым безусловным раздражителем. Когда затем у первых двух собак после довольно длинного периода с условными кислотными рефлексами условные раздражители еще раз подвергались превращению опять в пищевые, это превращение произошло очень быстро, через небольшое число раз. Значит, первая связь до известной степени уцелела, несмотря на существование второй связи.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Лекции о работе больших полушарий головного мозга»
Представляем Вашему вниманию похожие книги на «Лекции о работе больших полушарий головного мозга» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.



![Андрей Ангелов - Covid головного мозга [ноябрь]](/books/433274/andrej-angelov-covid-golovnogo-mozga-noyabr-thumb.webp)







Обсуждение, отзывы о книге «Лекции о работе больших полушарий головного мозга» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.