Александр Смирнов - Фармакология взаимодействия регуляторных пептидных систем головного мозга в механизмах подкрепления
Здесь есть возможность читать онлайн «Александр Смирнов - Фармакология взаимодействия регуляторных пептидных систем головного мозга в механизмах подкрепления» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Год выпуска: 2020, Жанр: Прочая научная литература, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Фармакология взаимодействия регуляторных пептидных систем головного мозга в механизмах подкрепления
- Автор:
- Жанр:
- Год:2020
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Фармакология взаимодействия регуляторных пептидных систем головного мозга в механизмах подкрепления: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Фармакология взаимодействия регуляторных пептидных систем головного мозга в механизмах подкрепления»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Фармакология взаимодействия регуляторных пептидных систем головного мозга в механизмах подкрепления — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Фармакология взаимодействия регуляторных пептидных систем головного мозга в механизмах подкрепления», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Вскоре после исследования грелина, как первого эндогенного лиганда для GHS-R1A (Kojima et al., 1999), стало ясно что эти рецепторы также являются потенциальной цель для контроля за приемом пищи и ожирением. На грызунах инъекции грелина (периферически или центрально) вызывали быстрый орексиновый ответ (Asakawa et al., 2001; Wren et al., 2000). Также было найдено, что хроническая стимуляция рецепторов грелином (Tschop et al., 2000) или синтетически выращенными гормонами (Lall et al., 2001) повышала массу тела у грызунов. Уровни грелина повышаются перед приемом пищи (Cummings et al., 2001) и была показана корреляция со степенью голода у здоровых особей (Cummings et al., 2004) определяющяя то, что, по крайней мере в нормальной физиологии, острые изменения грелина, как изменяющегося гормона голода, могут участвовать в формировании решения к употреблению пищи.
В экспериментальных исследованиях на грызунах были определены возможные зоны действия грелина в ГМ: дугообразное ядро, вентромедиальное ядро, дорсомедиальное ядро, паравентрикулярное ядро, латеральный гипоталамус (Olszewski et al., 2003 a,b; Wren et al., 2001 a,b), ядра одиночного тракта ствола мозга (Faulconbridge et al., 2003), центральное ядро миндалины (Olszewski et al., 2003), вентральная область покрышки (VTA) и прилежащее ядро (Egecioglu et al., 2010; Naleid et al., 2005).
Изучая механизмы награды, как ключевые цели для грелина, становится ясно что центральная система грелина необходима для развития наркотической зависимости (Jerlhag et al., 2009; Kaurand, Ryabinin, 2010; Tessari et al.,2007; Wellman et al.,2005 ), а грелиновый рецептор, GHS-R1A, является важной терапевтической мишенью при аддиктивном поведении.
Мезолимбические дофаминовые пути из вентральной области покрышки в прилежащее ядро, которые играют важную роль в побудительной мотивации (желании) (Berridge and Robinson, 2003), являются важным компонентом грелин-опосредованной системы. Грелиновые рецепторы, представленные в вентральной области покрышки (Guan et al., 1997; Zigman et al., 2006), включают субпопуляцию дофаминовых клеток в этой области (Abizaid et al., 2006). Воздействие грелина на вентральную область покрышки активирует и повышает высвобождение дофамина, а также активирует локомоторную стимуляцию (Jerlhag et al., 2006a, 2007) и снижение захвата дофамина в N.Acc. (Abizaid et al., 2006). Периферически введенный грелин также стимулирует мезолимбическую дофаминовую систему (Jerlhag, 2008; Quartaetal., 2009).
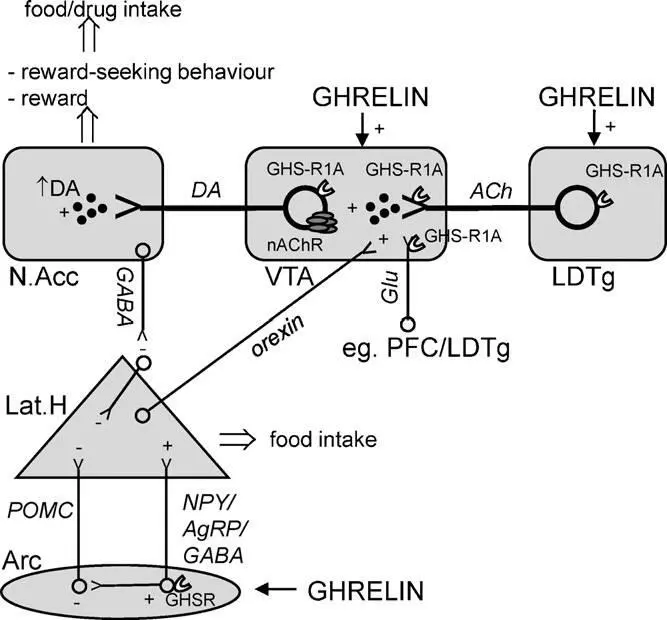
Схематичное изображение ключевых путей, через которые грелин регулирует пищевое поведение и поиск награды
Считается что грелин может иметь прямое влияние на узел подкрепления в LDTg (Guan et al., 1997) через GHS-R1A рецепторы, особенно выраженные на холинергических клетках (Dickson et all., 2010). Мы заметили, что локальное воздействие грелином в LDTg активирует показатели связанные с подкреплением, а именно локомоторную стимуляцию и высвобождение дофамина (Jerlhag et al., 2007). Более того, эти эффекты грелина были блокированы периферическим или интра-VTA воздействием неселективного холинергического никотинового антагониста, мекамиламином (Jerlhag et al., 2006a, 2008). К тому же, периферическая инъекция мекамиламина показала возможность блокирования грелина, локально воздействуя в VTA, чтобы повысить потребление пищи (Dickson et al., 2010). Совместно, эти исследования демонстрируют что грелин активирует холинергическо-дофаминергическую цепочку подкрепления, и таким образом может учавствовать в механизмах подкрепления.
В дальнейшем, с использованием селективного фармакологического антагониста субтипов никотиновых ацетилхолиновых рецепторов было показана способность грелина активировать холинергическо-дофаминергическую цепочку подкрепления опосредовано через особые субтипы никотиновых ацетилхолиновых рецепторов, а именно a3B2*, a6* и В3* (Jerlhag et al., 2008). Интересно, что эти субтипы также опосредуют подкрепляющие способности алкоголя и собственно потребление алкоголя грызунами (Ericson et al., 2009; Jerlhag et al., 2006b; Larsson and Engel, 2004; Lof et al., 2007; Steensland et al., 2007). Эти данные доказываются клиническими исследованиями; таким образом, блокируя эти субтипы рецепторов, возможно снижение употребляемой дозы (McKee et al., 2009).
Кроме того, было показано, что чрезмерное потребление алкоголя в длительный период времени, вызывает выброс ацетилхолина в VTA, следующий за повышением уровня прилежащего дофамина, доказывая, что алкоголь, как и грелин, активирует холинергическо-дофаминергическую цепочку подкрепления (Larsson et al., 2005). Анализирую полученные данные, возникают нейрохимические аналогии между грелином и алкоголем, где холинергическо-дофаминергическая цепочка подкрепления является главным регулятором.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Фармакология взаимодействия регуляторных пептидных систем головного мозга в механизмах подкрепления»
Представляем Вашему вниманию похожие книги на «Фармакология взаимодействия регуляторных пептидных систем головного мозга в механизмах подкрепления» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.




![Андрей Ангелов - Covid головного мозга [ноябрь]](/books/433274/andrej-angelov-covid-golovnogo-mozga-noyabr-thumb.webp)






Обсуждение, отзывы о книге «Фармакология взаимодействия регуляторных пептидных систем головного мозга в механизмах подкрепления» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.