К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
Здесь есть возможность читать онлайн «К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: Медицина, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
В течение многих лет было невозможно провести различие между потенциалами и теми пейсмекерами, которые связаны с явлением нарастания и убывания нейроэлектрических состояний, возникающих в микроструктуре соединений. Но, как уже было отмечено, Соколов (1970) показал, что в определенных лабораторных условиях механизм генерации аксонных нервных импульсов может действовать относительно независимо от изменений медленных потенциалов мембраны тела клетки. Согласно этим данным, генераторный потенциал возникает внутри структуры тела нейрона, а не на его поверхности и, следовательно, он не может быть отнесен непосредственно за счет изменений характеристик мембранного потенциала нервной клетки и дендритных влияний на них (см. также рис. IV-8).
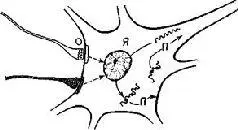
Рис. IV-8. Модель внутриклеточного контроля нейронных ритмов. Я – ядро; П – полипептид или протеин. Волнистая линия изображает переносчик (информационный) РНК, которая достигла или не достигла рибосом (). Соединение между субсинаптической мембраной, ядром и мембраной тела клетки, или аксоплазмой, показано сплошными стрелками. Синаптические пузырьки (О) показаны на нервном окончании вверху слева (см. Strumwasser, 1967).
Другая интересная диссоциация между генерацией импульсной активности и флуктуациями электрического состояния наблюдается во время одной из фаз сна. Эта фаза характеризуется быстрыми движениями глаз, часто подергивающимися движениями тела (ото можно увидеть, если понаблюдать за спящей собакой) и электрическими ритмами, которые могут быть записаны от мозга и которые неожиданно сходны с ЭЭГ бодрствующего состояния. Будучи разбужен на этой стадии сна, человек почти всегда сообщает, что он видел сны, тогда как у людей, проснувшихся во время других фаз сна, такие сообщения бывают редки.
Во время бодрствования у большей части нейронов мозга кошки (Evarts, 1967) повторяющаяся регулярность в разрядах не наблюдается. Во время обычного сна эти же клетки разряжаются взрывами спайков с периодами относительной неактивности между ними. Эти интервалы примерно соответствуют флуктуациям, одновременно записываемым макроэлектродами. Однако во время парадоксальной фазы сна нейроны дают интенсивные разряды с интервалами, повторяющимися примерно четыре раза в секунду, но электрические записи с макроэлектродов (ЭЭГ) не отражают этот доминирующий ритм. Это объясняется тем, что, хотя нейроны, разряжаясь, группируют свои спайки в четко выраженную временную структуру, они по фазе разряжаются независимо друг от друга, их активность не коррелирует с активностью соседних нейронов. Следовательно, корреляция спайковой активности отдельных нейронов не зависит от самого механизма генерации нервных импульсов. Поэтому должен существовать специальный механизм, осуществляющий связь спайковых разрядов соседних нейронов. Скорее всего, функцию этого механизма эффективно выполняет микроструктура медленных потенциалов, возникающих на контактах между нейронами. Таким образом, маловероятно, что устойчивое состояние и дискретные нейронные процессы являются взаимосвязанными процессами и отражают друг друга. Правила преобразования, которые позволяют перекодировать один процесс в другой, должны быть определены эмпирически для каждого из тех условий, которые интересуют исследователей. На данном этапе исследования любые общие утверждения относительно типов таких преобразований являются, вероятно, преждевременными.
Такая потребность в эмпирической информации, сообщающей о конкретных примерах преобразования дискретного процесса в непрерывный и наоборот, не исключает возможности создания гипотез о типах преобразований, которые, вероятнее всего, характезируют обычные операции перекодирования. Такие гипотезы необходимы еще и потому, что они определяют направление дальнейших исследований. По определению они будут соответствовать только категории обратимых преобразований, поскольку необратимые операции условны и, следовательно, должны быть просто открыты в эксперименте. Практически любой психологический процесс, который может быть описан словами, мог бы быть нейронно закодирован посредством необратимой операции. Однако остается открытым вопрос, действительно ли это является механизмом, пригодным для объяснения любого конкретного случая.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии»
Представляем Вашему вниманию похожие книги на «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.











Обсуждение, отзывы о книге «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.