К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
Здесь есть возможность читать онлайн «К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: Медицина, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Таким образом, перекодирование оказывается чрезвычайно эффективной частью процесса памяти, для осуществления которого конструкция нервной системы, по-видимому, великолепно .приспособлена. Формы перекодирования, которые возможны в нервной системе, фактически безграничны. Тем не менее можно выделить несколько классов кодов.
Мы уже разделили коды на два класса: коды, предполагающие дискретные импульсы нервного разряда, и коды, предполагающие микроструктуру устойчивого состояния, создаваемую нервными соединениями.
Проблема импульсного кодирования не сводится к вопросу о присутствии или отсутствии сигнала; импульсный код может использовать множество различных параметров, которые характеризуют последовательности залпов.
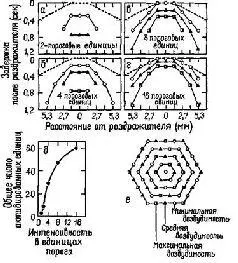
Рис. IV-4. Кодирование интенсивности и места раздражения, осуществляемое параллельными волокнами. Пространственное представительство показано в соответствии с тем, что было обнаружено в тактильных афферентах задней лапы кошки, когда последняя возбуждалась механическими залпами, приложенными к одной точке; диаграмма основывается на количественных экспериментальных данных. Если число равноотстоящих рецепторов с 3 уровнями возбудимости соответствует схеме е, то раздражитель интенсивностью в два порога вызывает некоторое число импульсов в стороне от раздражаемой точки и через какое-то время после раздражения а. Пунктирная линия обозначает латентный период, обусловленный временем распространения механической волны по лапе. Более сильные раздражители вызывают ответы, показанные на рис. б-г. Общее число импульсов в ответ на каждую интенсивность раздражителя представлено в д (Gray and Lai, 1965).
Д. Перкел и Т. Буллок (Perkel and Bullock, 1968), изучал сигналы, посылаемые электрическими органами некоторых рыб, установили следующие типы кодирования. При той или: иной комбинации условий изменяется длительность латентного периода, продолжительность спайковых взрывов, общая вероятность разрядов и вариация этой вероятности, увеличивается или уменьшается частота разрядов или скорость ее изменения. Такими условиями являются искажения электрического поля, создаваемые теми же самыми сигналами, посылаемыми электрическими органами, – искажения, регистрируемые системой боковой линии рыбы, этот процесс сходен с радарным.
Кроме этих организованных во времени кодов, имеет место и пространственное кодирование, когда множество параллельных линий – нервных волокон – продуцируют то, что Перкел и Буллой назвали обработкой «по ансамблю», которая зависит от различий в распределении последовательностей импульсов вдоль линий (см. рис. IV-4).
Пространственное кодирование и особенно кодирование, опирающееся на различия в синхронности разрядов у соседних групп нервов, тесно связано с целой группой процессов неимпульсного кодирования – микроструктурами медленных потенциалов, обеспечивающих устойчивое состояние, которые вносят существенный вклад в активность серого вещества мозга млекопитающих. Как говорят Перкел и Буллок, должно существовать какое-то считывающее устройство, какой-то механизм в центральной нервной системе, который считывает сообщения, закодированные в импульсной форме. В этих главах утверждается, что такое считывание возникает на местах соединений нейронов в процессе образования микроструктур медленных потенциалов, мгновенных состояний, возникающих в результате взаимодействия между совокупностями сигналов, действующих по соседству и кодируемых последовательностью импульсов.
Чтобы быть эффективными, большинство кодов нуждается в некотором стабильном уровне Работа Бернса и других, вне веяного сомнения, установила, что активность центральной нервной системы обладает такой стабильностью Нервная ткань спонтанно генерирует электрические потенциалы Мозг, как и сердце, непрерывно пульсирует И, так же как в сердце, такую пульсацию вызывают медленные потенциалы, а возникновение этих последних зависит от определенных констант химического окружения, в котором находится пульсирующая 1кань (рис IV-5).
Рис IV-5. Церебральная симфония (Verzeano et al, 1970).
Серии тщательно проведенных исследований в лаборатории Бернса (1958) дали исчерпывающий ответ на вопрос, который долгое время оставался фантастическим: может ли мозг сохранять активность даже в том случае, если он полностью изолирован (нейронально) от другой нервной ткани? Результаты этих экспериментов, как это часто бывает, не подтвердили полностью ни представления о том, что активность головного мозга «спонтанна», ни представления о мозге как находящейся в покое tabula rasa, на которую записывается сенсорный опыт. Берне обнаружил, что даже у неанестезированного животного изолированная полоска коры остается неактивной до тех пор, пока к ней хотя бы на короткое время не будет приложено электрическое раздражение; другие же данные (Echlin et al., 1952; Gerard and Joung, 1937; Henry and Scoville, 1952; Ingvar, 1955; Libet and Gerard, 1939) указывают на то, что спонтанная активность существует и в таких препаратах. В любом случае, даже если принять осторожный вывод, сделанный Бёрнсом, несколько сильных электрических раздражителей, приложенных к поверхности коры, вызывают серию разрядов нейронной активности, которая обычно продолжается в течение многих минут (или даже часов) после прекращения стимуляции.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии»
Представляем Вашему вниманию похожие книги на «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.











Обсуждение, отзывы о книге «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.