К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
Здесь есть возможность читать онлайн «К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: Медицина, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
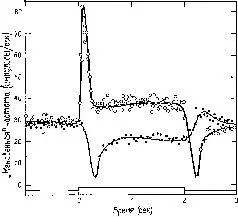
Рис. III-8. Одновременное кратковременное развитие возбуждения и торможения у двух соседних рецепторных единиц в латеральном глазе Limu-lus'a. Одна рецепторная единица (черные кружки) постоянно освещалась в течение всего периода, показанного на графике. Освещение другой единицы (белые кружки) также оставалось на одном постоянном уровне, кроме отрезка времени от 0 до 2 сек, когда ее освещение резко возрастало и оставалось на новом постоянном уровне. Заметный кратковременный рост возбуждения у одной рецепторной единицы сопровождается большими кратковременными тормозными эффектами у соседней, постоянно освещенной единицы. Резкое падение частоты вызывается тормозным эффектом, возникающим в результате большого кратковременного возбуждения. Во время постоянного освещения тормозный эффект еще присутствует, но менее заметен. Наконец, затухание частоты ответа у элемента, у которого уровень возбуждения был снижен, сопровождается заметным высвобождением от торможения (Ratliff, 1965).
Этот процесс «периферического», или «латерального», торможения не ограничивается зрительной системой. Сходную функциональную организацию имеют кохлеарный механизм слуха и рецепторы кожи (Bekesy, 1967). В центральной нервной системе аналогичным образом реагируют клетки коры головного мозга и коры мозжечка.
Короче говоря, латеральное торможение является одной нехарактерных особенностей нейронных сетей – особенно тех, которые организованы в плоские слои. В этих слоях располагается несколько уровней обработки информации о сигналу.
Какие же свойства этих нейронных сетей объясняют латеральное торможение? Было дано несколько объяснений латеральному торможению, и они не являются взаимно исключающими; точное математическое) описание и оценку каждого из них читатель может найти в обзоре Рэтлиффа (1965, гл. 3). Нуждается в объяснении прежде всего возникновение латерального торможения, его явная зависимость от расстояния до центра возбуждения, очевидность взаимодействия между возбуждением и торможением. Наиболее правдоподобное объяснение состоит в том, что разветвления каждого рецептора образуют тормозные связи с соседними рецепторами или, что более вероятно, они возбуждают тормозные нейроны – например, амакриновые и горизонтальные клетки сетчатки, обширные разветвления дендритов которых и отсутствие у них аксона делают их идеальными для выполнения этих функций (рис. III-9).
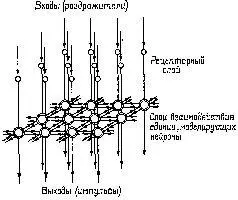
Рис. III-9. Схема рецепторного слоя и слоя взаимодействия (горизонтальные клетки) в сетчатке. Вертикальные стрелки указывают направление передачи сигнала, горизонтальные – тормозные взаимодействия (Ratliff, 1965).
Обнаружено, что такие не имеющие аксона дендритные сети в сетчатке (Svaetichin, 1967) и в каком-нибудь другом месте нервной системы (например, в мозжечке) выполняют точно такую же функцию. Действительно, недавние эксперименты (Werblin and Dowling, 1969) с использованием внутриклеточной регистрации показали, что горизонтальные клетки функционируют исключительно посредством гиперполяризации, то есть путем генерации тормозных медленных потенциалов. (Фактически от элементов сетчатки, расположенных на периферии слоя ганглиозных клеток, можно зарегистрировать только медленные потенциалы, а не нервные импульсы.) А возникновение торможения за счет дендритных сплетений, в которых медленные потенциалы генерируются в ответ на воздействия входных волокон, выявляя рисунок продуцируемых волновых форм, по существу, ведет к созданию эффектов интерференции, которые в их простейшей форме и образуют периферическое торможение.
В заключение следует отметить, что сенсорный механизм почти постоянно изменяет свою реакцию на входное воздействие, используя для этого два процесса: усиление контраста и адаптацию. Анатомические связи механизма делают вероятным реципрокное функционирование этих процессов; чем больше ослабевают ответы нейрона в ходе адаптации, тем меньшее тормозное влияние оказывает он на соседние нейроны, вызывающие усиление контраста (рис. III-10).
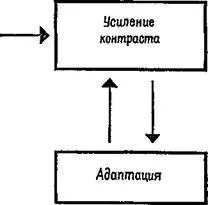
Рис. III-10. Реципрокные отношения усиления контраста и адаптации.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии»
Представляем Вашему вниманию похожие книги на «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.











Обсуждение, отзывы о книге «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.