К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
Здесь есть возможность читать онлайн «К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: Медицина, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Удивительно, что наши глаза находятся в постоянном движении – даже тогда, когда мы фиксируем точку. Эти небольшие, подобные тремору движения глаз можно зарегистрировать. Такие движения у некоторых людей настолько велики, что заметны другим, но – и в этом-то и состоит парадокс – человек с такими усиленными движениями глаз не знает о них до тех пор, пока не обратит на них внимание, когда смотрит на себя в зеркало (что обычно заставляет его обратиться к врачу, который, если он знаком с такой аномалией, успокаивает своего пациента, говоря, что это не опасно). Движения глаз препятствует тому, чтобы каждый из рецепторных элементов в течение какого-то отрезка времени возбуждался бы одним и тем же соотношением света и темноты, конечно, за исключением таких ситуаций, как плотный туман, когда свет теряет форму и функция зрения сводится к различению яркости. Чтобы изучить последствия нарушений таких движений глаз, на склере, белой части глазного яблока, не обладающей чувствительностью, укрепляют зеркало. Изображение проецируется на зеркало, отражается через призму на гладкую поверхность экрана, на которую смотрит наблюдатель. Призма корректирует отклонения рассматриваемого объекта, соответствующие отклонениям глазного яблока. Благодаря этому изображение, проецируемое на поверхность, всегда падает на одно и то же место сетчатки и образ стабилизируется (рис. Ш-3).
Стабилизированное изображение быстро становится незаметным. Зрительный прибор за несколько секунд так адаптируется, что изображение не может оставаться видимым – оно исчезает, адаптация завершена. Сходный опыт можно провести и в кожно-кинестетической системе: положите предмет на руку и некоторое время подержите его. Скоро ощущение наличия объекта исчезает.
Если бы не было такого механизма, организм подвергался бы непрерывной бомбардировке раздражителей разной длительности и интенсивности и это сделало бы его неспособным к тонкому различению. Фактически возможность зрительного различения, несмотря на изменение фонового освещения в диапазоне от 10 миллиардов до единицы, составляет одно из тех противоречий, которые вызывают большой поток исследований. В данном случае исследования адаптации сетчатки привели к открытию соответствующих нейронных механизмов. Это противоречие получило объяснение в концепции, согласно которой адаптация сетчатки целиком объясняется выцветанием и регенерацией фоточувствительного пигмента, содержащегося в рецепторах сетчатки (Hecht, 1934). Однако в последнее время стали накапливаться данные, говорящие о том, что для объяснения процесса адаптации необходимы и нефотохимические факторы я что эти факторы являются основой для осуществления фотохимических процессов (см. Rushton, в обзоре Dowling, 1967).
Главным инструментом в этих и‹ следованиях служит большой электрод, который помещают па внешней стороне глаза.
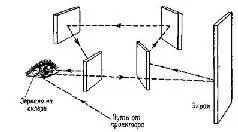
Рис. Ш-3. Благодаря тому, что стимул, прежде чем воздействовать на сетчатку, первоначально отражается от зеркала, укрепленного на склере и перемещающегося вместе с движениями глаз, последние могут быть компенсированы с такой точностью, что изображение раздражителя на сетчатке становится стабилизированным (см. Riggs et al, 1953).
Этот электрод записывает изменения потенциалов, генерируемых всей сетчаткой, в виде электроретинограммы. При соответствующих методах анализа получают такие же четкие ответы, как и тогда, когда используют микроэлектродную технику.
Коротко говоря, сетчатка млекопитающего состоит из нескольких слоев: рецепторный слой образован клетками, имеющими форму палочек и (или) колбочек, в которых имеются фоточувствительные химические пигменты; реагирующий первичный слой состоит из клеток, в которых процессы распространяются торизонтально и связывают друг с другом множество соседних клеток сетчатки; биполярный слой состоит из клеток, связывающих один или несколько рецепторов с ганглиозными клетками, являющимися началом выхода импульсов из сетчатки к мозгу ‹рис. Ш-4).
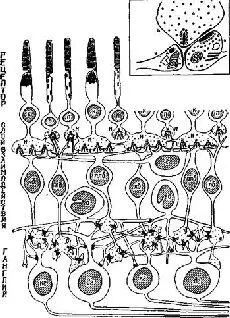
Рис. Ш-4. Схема строения сетчатки приматов, показывающая синапсы между различными типами клеток, наблюдаемые с помощью электронного микроскопа на серийных срезах. Обратите внимание на контакты между рецепторами, на широко распространенные связи горизонтальных и ама-криновых клеток и вертикальное расположение биполяров. Вздутые центральные окончания биполярных клеток формируют так называемые диад-ные синапсы (в рамке) с гаяглиозной клеткой и одновременно с амакри-новой клеткой. П – палочка; К – колбочка; кб – карликовый биполяр; по – палочковый биполяр; плб – плоский €иполяр: г – горизонтальная клетка; а – амакриновая клетка; кг – карликовая ганглиозная клетка; д – диадный синапс; дг – диффузная ганглиозная клетка; н – ножка. В прямоугольной рамке вверху справа показаны детали типичною диадного синапса (Horridge, 1968; Dowling and Boycott, 1966).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии»
Представляем Вашему вниманию похожие книги на «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.











Обсуждение, отзывы о книге «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.