К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
Здесь есть возможность читать онлайн «К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: Медицина, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
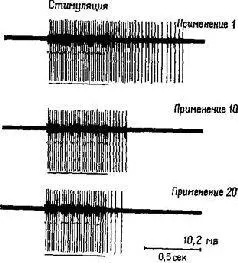
Рис. III-2, Затухающий ответ эфферентной единицы на повторяющееся электрическое раздражение кожи (Thompson, 1967).
Таким образом, в некоторых случаях процессы затухания, лежащие в основе привыкания, по-видимому, связаны не с пре- и постсинаптическими изменениями, а с какими-то другими механизмами. Однако в других случаях были получены противоположные результаты. Иногда \по не всегда) этот процесс связан с уменьшением или увеличением притока медиаторных веществ,
С. Шарплесс (1967, 1969) показал, что при стимуляции нейронов у позвоночных не возникает ни сенситизации, ни десенси-тизации их сияаптических мембран. Напротив, Э. Кендел (Castel-lucci et al., 1970; Kupfermann et al., 1970; Pinsker et al., 1970) нашел, что привыкание и его нарушение у беспозвоночных локализуются в пресинаптических окончаниях и предполагают участие эффективного механизма секреции возбуждающих веществ в синапсах.
Стало быть, полученные данные не согласуются между собой. Необходимо найти ответ на целый ряд вопросов. Каким образом вообще механизм возникновения возбуждений может испытывать влияния, если не через посредство медленных потенциалов? Запускается ли внутри нейрона некоторый биохимический процесс и если да, то как (особенно если мы имеем дело с системой, которая обычно обнаруживает спонтанную активность и, следовательно, каким-то образом генерирует потенциалы)? Какова природа биохимических изменений и как они развиваются во времени? Соколов предполагает, что в этом механизме участвует РНК. Если он прав, то не связано ли привыкание с первыми этапами индукционного процесса или механизм привыкания совершенно независим от процесса постоянного хранения следов памяти? Возможно, что механизм постоянного хранения действует только при условии длительного и повторного привыкания. Если это так, то как это происходит? Наконец, каким образом изменение в структуре нервного импульса, вызываемого привыканием, действует на микроструктуру медленного потенциала на следующем синаптическом уровне? Чтобы ответить на некоторые из зтих вопросов, рассмотрим нейронную организацию сетчатки, этого маленького «кусочка мозга», который является гораздо более доступным для изучения, чем центральная нервная система.
АДАПТАЦИЯ
То, что мы воспринимаем из нашего окружения, зависит от возможностей наших рецепторных органов, которые накладывают свои ограничения на восприятие той или другой конфигурации воздействия. Так, оптическая система глаза фокусирует на сетчатку ограниченный диапазон электромагнитных волн; звуковые волны оказывают давление на жидкость в улитке уха; разного рода деформации кожи возбуждают свободные или специализированные нервные окончания в соматической системе и т. д. Эти формы энергетических изменений взаимодействуют « собственной активностью рецепторов и вызывают ее изменения, которые являются достаточно надежными, чтобы организм мог их идентифицировать как результат внешних воздействий.
Воспользуемся еще раз простой моделью протекания речевого акта, описанного в гл. I. Там участвовали две основные переменные: устойчивое состояние и дискретные переменные. Эта модель была применена к функции мозга, сейчас мы используем ее для сенсорного механизма. Заменим состояние постоянного напряжения голосовой связки постоянным распределением различных видов энергии по рецепторным поверхностям, дискретные колебания воздуха – дискретными нейронными разрядами в форме нервных импульсов. Короче говоря, предположим, что то, что происходит в органе чувств, совсем не отличается от того, что происходит в нервной системе, и что нейрофизиология сенсорных процессов может служить миниатюрной моделью ориентировочного рефлекса и процесса привыкания.
Всем известен процесс адаптации: ощущение при погружении в слишком горячую ванну и осознание спустя несколько минут, что для полного удовольствия следовало бы добавить еще теплой воды; исчезновение ощущения давления на кожу, ожидание в течение нескольких минут после входа в темный зал кинотеатра,, прежде чем мы что-либо увидим и будем способны найти свободное место. Можно привести много примеров сенсорной адаптации. Наиболее яркий из них тоже парадоксален и был получен в экспериментах, при которых проецируемый на сетчатку образ стабилизировался с помощью зеркал и линз (Ditchburn and Ginsborg, 1952; Riggs, Rattliff, Cornsweet and Comsweet, 1953).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии»
Представляем Вашему вниманию похожие книги на «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.











Обсуждение, отзывы о книге «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.