К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
Здесь есть возможность читать онлайн «К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: Медицина, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Вначале думали, что моторная система уникальна и центральный контроль над рецепторами существует только в двигательной системе. Однако очень скоро стали накапливаться данные о том, что кожные рецепторы (Hagbarth and Kerr, 1954), слуховой афферентный механизм (Galambos, 1956; Rasmussen, 1946; Desmedt r 1960; Dewson, 1968) и обонятельная чувствительность (Kerr and Hagbarth, 1955) также являются объектом прямой регуляции со‹ стороны центральной нервной системы. Не сразу удалось доказать существование эфферентного контроля зрительного входа; первые исследования (Granit, 1955; Hernandez-Peon and Scherer 1955) были подвергнуты критике потому, что еще не найдены морфологически эфферентные волокна в сетчатке. Поэтому в моей лаборатории были проведены исследования в надежде, что эфферентный контроль над зрительным входом станет вполне реальным фактом. В этих исследованиях были использованы только что появившиеся компьютеры и микроэлектродная техника. Результаты опытов показали, что незрительные стимулы (звуковые щелчки и удары по лапе) вызывают ответы в зрительном нерве у кошек (мышцы которых, в том числе и мышцы зрачка, были обездвижены). Кроме того, электрическая активность -сетчатки (измеряемая с помощью электроретинограммы) и аффе-рентов, берущих начало в сетчатке, также меняется в результате такой незрительной стимуляции (Spinelli, Pribram and Weingar-ten, рис. V-3).

Рис. V-3. Запись электрических ответов на звуковой щелчок с помощью биполярных электродов, вживленных в зрительный нерв. Запись осуществлена непосредственно на фотопленку с осциллоскопа методом суперпозиции. Левая запись была сделана, когда животное обнаруживало реакцию внимания, а правая – во время его отвлечения (Spinelli, Pribram and Weingarten, 1965).
Наконец, как и в случае других сенсорных модальностей, стимуляция соответствующей части коры головного мозга вызывала изменение в рецептивных полях ганглиозных клеток сетчатки, например их величины (Spinelli and Pribram, 1967). Результаты этих экспериментов убеждают в том, что организация зрительного механизма сходна с организацией других сенсорных систем, в которых существует центральный контроль над сенсорным выходом.
Один процесс, участвующий в нейронном контроле над входом, отличается от до сих пор рассмотренных обратных связей. Интерес к нему возник после того, как великий немецкий физик и психофизиолог Герман Гельмгольц обнаружил некий парадокс в области восприятия. Он заметил, что зрительное изображение мира начинает прыгать, когда мы пальцем надавливаем на наше глазное яблоко. Напротив, при произвольном движении глаз или при движении ими в ответ на внешние стимулы зрительное изображение мира остается неподвижным. В обоих случаях при манипулятивном и обычном движении глаз зрительные структуры, проецирующиеся на сетчатку, идентичны. Гельмгольц сделал вывод, что обычный процесс восприятия должен, следовательно, включать некий механизм, рассчитывающий и корригирующий сигналы, поступающие от сетчатки, насколько это необходимо для получения неискаженного образа.
Мертон (Brindley and Merton, 1960) проверил наблюдения Гельмгольца экспериментально. Он парализовал мышцы своего собственного глаза, а затем попытался двигать глазами. Оказалось, что изображение зрительного мира прыгало в том же направлении, в каком он пытался переместить свой взор.
Было бы логично предположить, что в осуществлении коррекции, необходимой для обычного восприятия мира, принимает участие обратная связь от мышц глаза. Однако результаты экспериментов с параличом мышц глаза плохо согласуются с этой гипотезой, поскольку движение изображения зрительного мира не связано ни с какими сокращениями мышц. Кроме того, пути, проводящие проприоцептивные импульсы от мышц глаза, не были прослежены в мозгу, несмотря на многочисленные попытки это сделать. Все эти факты заставляют предположить, что существует какой-то «опережающий» процесс, который корректирует восприятие (МасКау, 1966; Mittelstaedt, 1968).
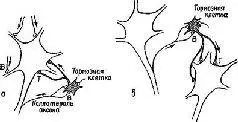
Рис. V-4. Затухание нейронной активности вследствие действия отрицательной обратной связи, осуществляемой клетками Реншоу (а). Усиление контраста посредством активации клеток Реншоу, вызывающих торможение у соседних нейронов (б). Это сужает центральное поле разряжающихся спайками клеток и создает в нейронной сети чередующиеся полосы возбуждения и торможения.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии»
Представляем Вашему вниманию похожие книги на «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.











Обсуждение, отзывы о книге «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.