Ирина Спивак - Репликация ДНК - учебное пособие
Здесь есть возможность читать онлайн «Ирина Спивак - Репликация ДНК - учебное пособие» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: СПб., Год выпуска: 2011, Жанр: Химия, Биология, Биология, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Репликация ДНК: учебное пособие
- Автор:
- Жанр:
- Год:2011
- Город:СПб.
- ISBN:нет данных
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Репликация ДНК: учебное пособие: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Репликация ДНК: учебное пособие»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Репликация ДНК: учебное пособие — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Репликация ДНК: учебное пособие», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Различия в продолжительности S-фазы найдены и у других организмов. Например, у тритона S-фаза длится 1 ч в ядрах бластулы и 200 ч в предмейотической S-фазе сперматоцитов. Вероятно, длительность S-фазы определяется не скоростью синтеза ДНК, а числом задействованных ориджинов репликации. В ДНК клеток нейрулы тритона они находятся на расстоянии около 40 мкм друг от друга, а в соматических клетках – около 100 мкм.
Таблица 1
Число и длина репликонов у разных организмов.

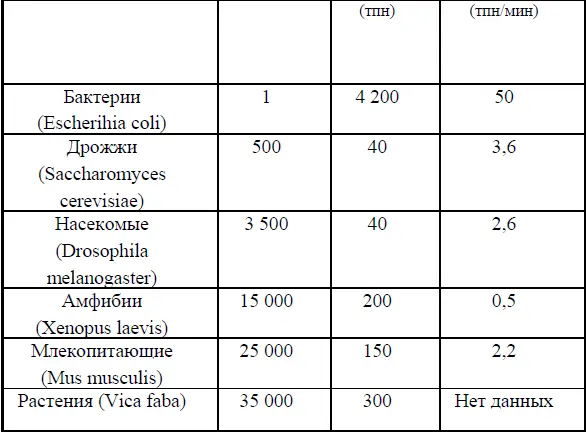
В соответствии с современными представлениями репликоны у эукариот распределены в геноме не случайно, они расположены группами (replicon foci). В этих группах, или фокусах, собираются ферменты репликации, которые удлиняют вилки репликации одновременно 10-100 соседних репликонов длиной примерно по 100тпн каждый. Репликация в них завершается за 45–60 мин. Кроме этого существуют очень длинные репликоны (более 1000тпн) – столь большие, что репликация в них продолжается по нескольку часов.
Активация ориджинов репликации происходит на протяжении всей S-фазы. Например, АRS1 S.cerevisiae активируется в ранней, а АRS501 – в поздней S-фазе. Большинство ориджинов активируется в середине S-фазы. Интересно отметить, что участки хромосом S.cerevisiae, реплицирующиеся в ранней или поздней S-фазе, располагаются мозаично, то есть перемешаны. У S.cerevisiae было обнаружено, что центральная область хромосомы IV реплицируется в ранней, а теломеры – в поздней S-фазе. Участок ДНК размером 67тпн, прилежащий к теломере на правом конце хромосомы V и содержащий АRS501, реплицируется в поздней S-фазе. По-видимому, поздняя репликация этого участка хромосомы является следствием его соседства с теломерой. Кроме того, известно, что в конце S-фазы реплицируются "молчащие" гены, например неэкспрессирующиеся в определенных типах клеток локусы НМL и НМR, которые локализованы в субтеломерных областях. Активно экспрессирующиеся гены, например локус МАТ, напротив, реплицируются в первой половине S-фазы.
Глава 3. Инициация репликации
Ориджины репликации являются местом, с которого начинает свое движение репликативная вилка. Но ДНК-полимеразы не могут начать процесс репликации без помощи других белков. Белки, участвующие в распознавании ориджина и спсобствующие привлечению к нему праймазы – РНК-полимеразы, синтезирующей праймер, «затравку» для синтеза ДНК – и ДНК-полимеразы, образуют комплекс инициации репликации.
3.1 Инициация репликации у E.coli
Инициация репликации в оriС в системе in vitro начинается с формирования комплекса, в состав которого входят шесть белков: DnaА, DnaВ, DnaС, НU, Girase и SSВ. Сначала с девятичленной последовательностью связывается мономер DnaА, затем 20–40 мономеров этого белка формируют большой агрегат. ДНК ориджина опоясывает его, и цепи ДНК разъединяются в области трех тринадцатичленных последовательностей. На следующем этапе димер DnaВ/DпаС присоединяется к комплексу oriС/DnaА, формируя агрегат размером около 480 кДа, соответствующий сфере с радиусом 6 нм. В результате формируется вилка репликации.
3.2. Инициация репликации у эукариот
Инициация репликации ДНК эукариот начинается с образования комплекса ориджина репликации и белка-инициатора репликации. Этот комплекс называется пострепликативным (роst.-RС). Он служит платформой для сборки структур более высокого порядка, которые переводят хроматин в состояние, компетентное для репликации. Последовательные стадии образования комплексов инициации репликации показаны на рис. 7.
Белком-инициатором репликации ДНК в клетках эукариот является ОRС (origin recognition complex), который впервые был описан у S.cerevisiae. Впоследствии ORC-подобные белки были обнаружены и изучены и у других представителей эукариот, а также у млекопитающих и человека. У всех эукариот ОRС образован шестью субъединицами – Огс1-Огсб (120-50 кДа). Для жизнедеятельности S. cerevisiae существенны все шесть субъединиц комплекса. Две разные группы субъединиц ОRС участвуют в распознавании последовательностей ориджина при его связывании с ориджином репликации. Огс1, Огс2 и Огс4 взаимодействуют с АСS, остальные три субъединицы распознают B1– подобные элементы. Возможно, что связь с нуклеотидными последовательностями В1 осуществляет только Огс5. ОRС специфически связывается с ДНК только в присутствии АТР и обладает АТР-азной активностью, которая регулируется координированным взаимодействием белка с АТР и элементами АRS. АТР связывается с субъединицей Огс1 и играет роль кофактора, необходимого для присоединения ОRС к ориджину. Специфическая последовательность ориджина, связавшегося с ОRС, ингибирует АТРазную активность Огс1, в то время как однонитевые участки ДНК, появляющиеся в S-фазе, ее снова активируют. При этом меняется конформация ОRС – с вытянутой (ехtended) на изогнутую (bеnt). Возможно, связывание и гидролиз АТР субъединицей Огс1 участвуют в контролировании функций ОRС в клеточном цикле.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Репликация ДНК: учебное пособие»
Представляем Вашему вниманию похожие книги на «Репликация ДНК: учебное пособие» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Репликация ДНК: учебное пособие» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.