Ирина Спивак - Репликация ДНК - учебное пособие
Здесь есть возможность читать онлайн «Ирина Спивак - Репликация ДНК - учебное пособие» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: СПб., Год выпуска: 2011, Жанр: Химия, Биология, Биология, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Репликация ДНК: учебное пособие
- Автор:
- Жанр:
- Год:2011
- Город:СПб.
- ISBN:нет данных
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Репликация ДНК: учебное пособие: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Репликация ДНК: учебное пособие»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Репликация ДНК: учебное пособие — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Репликация ДНК: учебное пособие», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Уотсон и Крик уже во второй своей работе 1953 г. предположили возможный механизм копирования наследственного материала. Легко представить, что цепи молекулы ДНК расходятся и каждая из них становится матрицей, на которой синтезируется новая комплементарная цепь. В результате образуются две дочерние двуспиральные молекулы ДНК, не отличимые от родительской молекулы.
В 1957 г. А. Корнберг обнаружил у бактерии Е. соli фермент, катализирующий процесс полимеризации ДНК из нуклеотидов – ДНК-полимеразу 1. В 1959 г. Артуру Корнбергу (А. Kornberg) была присуждена Нобелевская премия за открытие механизма биосинтеза ДНК. Он показал, что в основе удвоения молекул ДНК лежат обычные биохимические реакции.
В общем виде реакцию присоединения 5'-дезоксинуклеотидной группы к З'-ОН-группе концевого нуклеотида праймерной цепи можно представить следующим образом:
[dNMP] n+ dNTP ↔ [dNMP] n+1+ РР i
где dNMP– любой из четырех обычных нуклеотидов. За один акт репликации нить, содержащая 3’-конец, удлиняется на один нуклеотидный остаток, при этом одновременно происходит удаление пирофосфата. Реакция присоединения нуклеотида обратима, но так как неорганический фосфат в клетках быстро разрушается, то реакция активно направлена в сторону синтеза. Репликация ДНК всегда идет от 5’– конца нити ДНК (то есть содержащего 5’-дезоксинуклеотидную группу) к 3’-концу (то содержащему свободную 3-ОН-группу) и нуждается в наличии ранее синтезированного фрагмета нити ДНК в качестве затравки для реакции полимеризации. Такой ДНК-фрагмент, имеющий свободный 3’-конец, называется праймером. Ферменты, катализирующие праймер-зависимую, детерминируемую ДНК-матрицей реакцию присоединения дезоксинуклеотидов, называются ДНК-полимеразами. К настоящему времени выделены и охарактеризованы несколько различных классов ДНК-полимераз, детально описаны свойства этих ферментов и реакции, которые они катализируют. Об их строении и индивидуальных особенностях мы подробно поговорим в следующих главах.
1.1. Вилка репликации
Процесс репликации происходит в специальных структурах, названных вилками репликации. Схематическое устройство репликативной вилки E.coli представлено на рис. 1. То, что две нити молекулы ДНК расположены антипараллельно друг другу, создает ряд проблем для их олдновременной разнонаправленной репликации.
По мере движения вилки одновременно должны синтезироваться две дочерние цепи. Вилка движется в направлении от 5 ' к 3’ на одной цепи и от 3 ’ к 5' – на другой. Однако нуклеиновые кислоты синтезируются только от 5'– к 3'-концу. Проблема решается таким образом, что на одной из родительских нитей новая нить синтезируется непрерывно в направлении 5'-3', что совпадает с движением вилки репликации. Это называется лидирующей или ведущей. Другая нить называется отстающей или запаздывающей, так как синтез на ней идет с некоторой задержкой по сравнению с лидирующей нитью. Это связано с тем, что ДНК на этой нити синтезируется также от 5' к 3', но в направлении, противоположном движению вилки, и короткими фрагментами. Благодаря этому разнонаправленный синтез ДНК может осуществляться в рамках одной структуры – репликативной вилки.
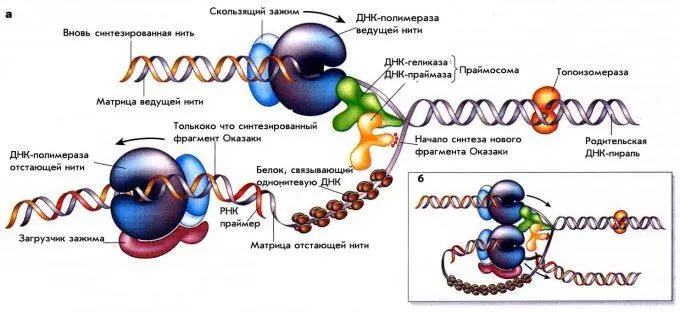
Рис. 1. Схема репликативной вилки.
Длина таких коротких фрагментов у прокариот составляет 1000–2000 пн. По имени открывшего их ученого они были названы «фрагментами Оказаки». По мере движения репликативной вилки концы соседних фрагментов Оказаки соединяются с образованием непрерывной отстающей нити. Для того, чтобы процесс на обеих нитях шел синхронно, полимеразные комплексы лидирующей и отстающей нити связаны между собой, образуя сложную трехмерную структуру (рис. 1, б)
Вилка репликации может двигаться как в одну сторону от точки начала репликации, так и в обе стороны. В зависимости от этого процесс называется однонаправленной или двунаправленной репликацией. Как это выглядит схематически, показано на рис. 2. У эукариот репликация обычно двунапраленная. Также и у E.coli .
Механизмы инициации репликации в точке начала репликации и при образовании фрагментов Оказаки в отстающей цепи в принципе аналогичны, хотя имеются некоторые тонкие различия. В обоих случаях происходит образование коротких РНК-затравок (праймеров ),комплементарных матричной ДНК, в виде продолжения которых синтезируется новая цепь ДНК. В дальнейшем короткие вставки РНК замещаются сегментами ДНК, отдельные фрагменты Оказаки затем объединяются с образованием непрерывной отстающей нити.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Репликация ДНК: учебное пособие»
Представляем Вашему вниманию похожие книги на «Репликация ДНК: учебное пособие» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Репликация ДНК: учебное пособие» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.