Ирина Спивак - Репликация ДНК - учебное пособие
Здесь есть возможность читать онлайн «Ирина Спивак - Репликация ДНК - учебное пособие» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: СПб., Год выпуска: 2011, Жанр: Химия, Биология, Биология, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Репликация ДНК: учебное пособие
- Автор:
- Жанр:
- Год:2011
- Город:СПб.
- ISBN:нет данных
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Репликация ДНК: учебное пособие: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Репликация ДНК: учебное пособие»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Репликация ДНК: учебное пособие — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Репликация ДНК: учебное пособие», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Все живые организмы на Земле обычно делят на прокариот и эукариот (от греч. карион – ядро). Главной особенностью прокариот является отсутствие у них в отличие от эукариот полноценного клеточного ядра, покрытого оболочкой. Генетический материал прокариот расположен в нуклеоиде – примитивном эквиваленте ядра эукариот. Клетки прокариот имеют очень небольшие размеры – около 1 мкм. Объем эукариотических клеток в 800-1000 раз больше объема клеток прокариот. К прокариотам относятся бактерии и археи (или архебактерии), предки которых возникли около 4 млрд лет назад. Эукариоты могут быть как одноклеточными, так и многоклеточными. Они появились на Земле примерно через 500 млн лет после прокариот.
По современным представлениям ДНК-метаболизм у прокариот имеет некоторые отличия от такового у эукариот. Описывая процессы репликации и рекомбинации, мы будем каждый раз подчеркивать эти отличия.
Глава 2. Начало репликации
Репликация ДНК начинается не в любой случайной точке молекулы, а в специфических местах, называемых точками начала репликации или олриджинами. Процесс копирования продолжается через образование репликативных вилок в одном или обоих направлениях до тех пор, пока ДНК полностью не удвоится. В замкнутых кольцевых молекулах ДНК новосинтезированные цепи ковалентно соединяются в местах встречи увеличивающихся в размере репликативных вилок или в том месте, где единственная вилка возвращается к точке начала репликации. Дочерние молекулы, как правило, расходятся еще до начала нового раунда репликации.
Такие различающиеся по размеру геномы, как геном вируса SV40 (5,2тпн), бактериофага λ (48,5тпн) и Е. соli (4-10 3тпн), воспроизводятся в результате одного инициирующего события, происходящего в определенной точке.
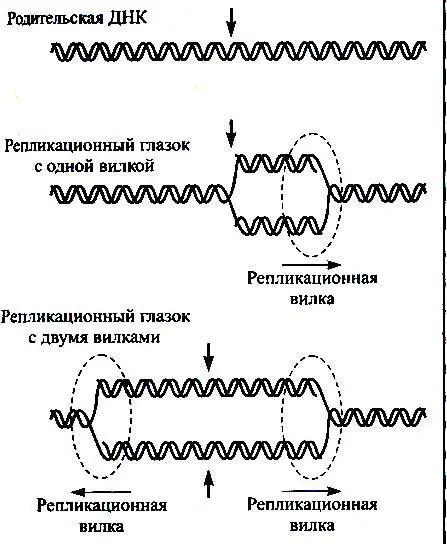
Рис. 2. Возможное движение репликативной вилки.
У про– и эукариот можно встретить различные вариации на эту тему. Так, каждая из цепей родительской спирали митохондриальной ДНК животных (15тпн) имеет свою точку начала репликации. Синтез комплементарной цепи некоторых небольших однонитевых фаговых геномов начинается вблизи одной специфической последовательности, а репликация полученного дуплекса может инициироваться совсем в другой точке. Репликация линейных двунитевых ДНК также инициируется в особых сайтах. Например, ДНК бактериофага Т7 (40тпн) реплицируется в двух противоположных направлениях к разным концам молекулы, начиная от одной точки, а каждая из двух цепей ДНК аденовируса человека (30–38 тпн) реплицируется последовательно всегда от З'-конца.
Для геномов эукариотических клеток характерно наличие множественных точек начала репликации, разбросанных по хромосоме на расстоянии около 20тпн. После инициации репликация продолжается в двух направлениях от каждой точки до тех пор, пока репликативные вилки двух соседних точек начала репликации не сольются. Полноразмерные ДНК каждой дочерней хромосомы получаются путем соединения более коротких, независимо инициированных новосинтезированных нитей.
2.1. Понятие о репликоне и ориджине репликации
Участок ДНК, на котором синтезируется отдельный фрагмент лидирующей нити, называется репликоном. У многих прокариот их геном содержит только одну точку инициации репликации, то есть у них в ДНК только один репликон. Эукариотические геномы полирепликонны.
Место начала репликона, в котором происходит инициация репликации, носит название ориджина репликации. Именно ориджин распознается специальными белковыми комплексами и на нем начинается формирование вилки репликации.
В некоторых случаях место начала репликации имеет такую нуклеотидную последовательность, что дуплекс принимает необычную конфигурацию, которую распознают белки, участвующие в инициации. Природа взаимодействия между точкой начала репликации и белками и механизм инициации в целом исследованы недостаточно, однако можно сказать, что, по-видимому, они в разных случаях различны.
2.2. Ориждин репликации E.coIi oriC
Наиболее подробно изучены ориджины у Е. соli и Bacillus subtilis. Область начала репликации хромосомы, оriС (origin of chromosome), включает в себя участки со специфическими последовательностями, так называемыми ДНК-боксами, и расположенными между ними короткими последовательностями. ДНК-боксы со специфическим «мотивом» нуклеотидов, преимущественно в 9пн, перемежаются фрагментами в 12-1Зпн с высоким содержанием АТ. Сами девятичленные последовательности могут располагаться как в прямом, так и в инвертированном положении по отношению друг к другу. Например, у В. subtilis имеется один фрагмент ТТАТССАСА и два других девятичленных бокса, ориентированных в противоположном направлении, с заменой одной из пар нуклеотидов. Всего у В. subtilis на оriС расположено 15 ДНК-боксов. Область оriС очень консервативна: ДНК-боксы сходного состава имеются в соответствующем месте хромосомы у других бактерий (только у Mycoplаsma genitalium, несмотря на наличие общих для всех бактерий ферментов репликации, ДНК-боксов найдено не было). Сами ДНК-боксы не кодируют белок или РНК, хотя между ними располагаются отдельные гены. Продукты этих генов также большей частью вовлечены в «обслуживание» процесса репликации ДНК.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Репликация ДНК: учебное пособие»
Представляем Вашему вниманию похожие книги на «Репликация ДНК: учебное пособие» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Репликация ДНК: учебное пособие» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.