Феликс Филатов - Клеймо создателя
Здесь есть возможность читать онлайн «Феликс Филатов - Клеймо создателя» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. ISBN: , Издательство: Array Литагент «Ридеро», Жанр: Биология, Прочая научная литература, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Клеймо создателя
- Автор:
- Издательство:Array Литагент «Ридеро»
- Жанр:
- Год:неизвестен
- ISBN:978-5-4474-2574-6
- Рейтинг книги:4.67 / 5. Голосов: 3
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Клеймо создателя: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Клеймо создателя»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Клеймо создателя — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Клеймо создателя», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Разделим по аналогии с АРСазами-1 и -2 – соответствующие им аминокислоты также на два класса арс-1 и арс-2. При этом возникает внятная билатеральная симметрия двадцатки аминокислот: ровно половина из них (мы здесь не вдаемся в детали), синтезируется с помощью аминоацил-тРНК-синтетаз (АРСаз) I класса:
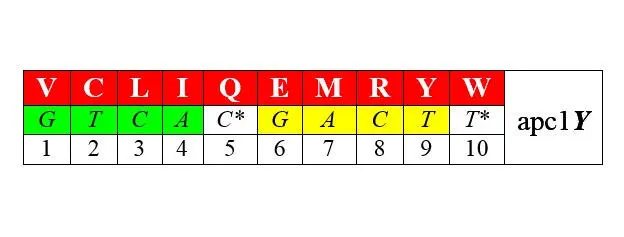
Другая половина синтезируется с помощью АРСаз II класса (нижние строки – порядковые номера аминокислот при раздельной – по классам [1—10 и 1—10] и при сплошной [1—20] их нумерации):
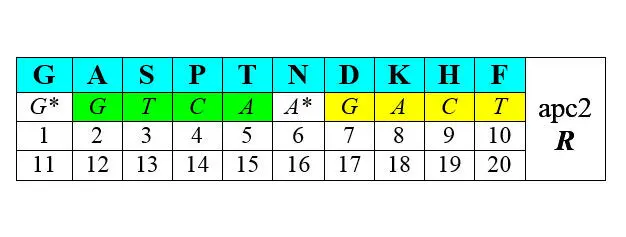
В обоих представленных рядах аминокислоты упорядочены по нарастанию молекулярной массы. Любопытно, что в каждый арс-класс попадает по три неполярные алифатические аминокислоты (V LIи GAP), по три полярные незаряженные ( CQMи STN), по одной негативно и позитивно заряженных ( E-R+и D-K+) и по две ароматические ( YWи HF) аминокислоты. В каждой из строк первых букв кодирующих эти аминокислоты триплетов легко различаются две четверки GTCA и GATC , разделенные в одном случае пиримидинами С* и Т* , в другом – пуринами G* и А* . Поэтому арс-1 мы условно называем пиримидиновыми аминокислотами арс- Y , а арс-2 – пуриновыми , арс- R . Как это может соотноситься с молекулярной биологией процессов, связанных с трансляцией, мы увидим далее.
Описанные симметрии рядов арс- Y и арс- R сохраняются и в двумерном (2D) представлении. Это представление, образно (то есть, не математически) названное Автором базовой матрицей генетического кода, формируется абсциссой, вдоль которой размещаются аминокислоты, упорядоченные по нарастанию молекулярных масс, и ординатой, вдоль которой размещаются первые кодирующие основания, соответствующие этим аминокислотам.
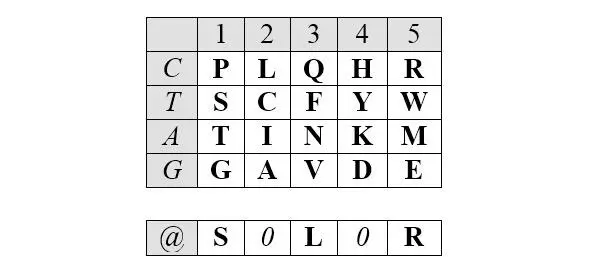
Базовая (без нижней дополнительной строки) матрица является прямоугольником 4х5, содержащим двадцатку канонических аминокислот. В отличие от каллигаммы, матрица не требует специального допущения для кодирования цистеина и, таким образом, полностью соответствует универсальному генетическому коду. Центральные колонки матрицы отчетливо структурированы по гидрофильности аминокислот (в таблице ниже – светлые ячейки заняты гидрофобными аминокислотами, темные – гидрофильными):
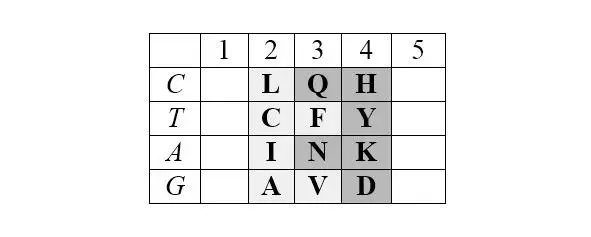
Матрица структурирована также по позициям обозначенных выше четверок аминокислот обоих арс-классов, демонстрируя их строгую сдвиговую симметрию:
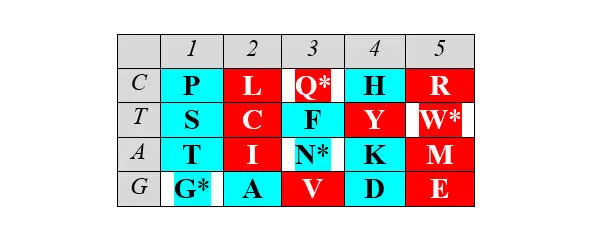
Отчетливо заметна и симметрия продуктов, маркирующих деление арс-классов аминокислот на «пуриновые» и «пиримидиновые», – относительно центральной колонки матрицы:
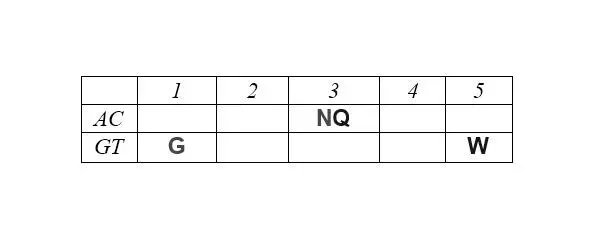
Поскольку в каждой строке матрицы не более двух или трех аминокислот одного класса с хорошо различимой массой, то при способности АРСаз узнавать хотя бы первое основание кодона (третье – антикодона) и отличать его от фиксированного по массе реперного соседа, безошибочное узнавание АРСазой «своей» тРНК чрезвычайно упрощается. Таким образом, матрица хорошо иллюстрирует решение парадокса множественного узнавания небольшой молекулы с почти монотонной структурой.
Заканчивая главу, упомянем еще об одной нашей находке, имеющей отношение к кодовым симметриям. Оказывается, продукты кодирования зеркально-симметричными дублетами (типа ABN и BAN ) следуют трем простым правилам:
при нарастании массы оснований в дублете кодируемый продукт имеет б о льшую молекулярную массу, при снижении – меньшую;
молекулярная масса продукта, кодируемого гомотриплетом (то есть, ССС , ТТТ , ААА , GGG ), больше, если триплет составлен из пиримидинов;
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Клеймо создателя»
Представляем Вашему вниманию похожие книги на «Клеймо создателя» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.









![Наталья Александрова - Клеймо сатаны [= Табакерка Робеспьера] [litres]](/books/404375/natalya-aleksandrova-klejmo-satany-tabakerka-ro-thumb.webp)

Обсуждение, отзывы о книге «Клеймо создателя» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.