Феликс Филатов - Клеймо создателя
Здесь есть возможность читать онлайн «Феликс Филатов - Клеймо создателя» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. ISBN: , Издательство: Array Литагент «Ридеро», Жанр: Биология, Прочая научная литература, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Клеймо создателя
- Автор:
- Издательство:Array Литагент «Ридеро»
- Жанр:
- Год:неизвестен
- ISBN:978-5-4474-2574-6
- Рейтинг книги:4.67 / 5. Голосов: 3
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Клеймо создателя: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Клеймо создателя»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Клеймо создателя — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Клеймо создателя», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
После транскрибирования мРНК не остается комплементарно связанной с ДНК-шаблоном; она освобождается от ДНК, которая затем восстанавливает свою двойную структуру. В одной эукаритической клетке количество молекул мРНК может составлять свыше 10.000. Наряду с молекулами мРНК на ДНК образуются и другие транскрипты, в том числе молекулы рибосомной и транспортных РНК, которые также имеют важное значение в реализации генетической информации. Все эти РНК называют еще ядерными. Наиболее обильными РНК в клетках всех видов являются молекулы рибосомной РНК (рРНК), которые выполняют роль структурных компонентов рибосом. У эукариот синтез рРНК контролируется огромным количеством генов (сотни и тысячи копий) и происходит в ядрышке. Не похоже, что структура рРНК имеет серьезное значение для обсуждаемой далее формальной организации генетического кодирования, и мы не станем на ней останавливаться. Совершенно другую роль в этих процессах имеют встречающиеся в клетке в несколько меньших количествах молекулы транспортных РНК (тРНК), которые участвуют в декодировании информации, трансляции. Это те самые декодеры или молекулы-посредники ( codemakers Барбьери), – которые обусловливают специфическую связь между хранилищем генетической информации, нуклеиновыми кислотами, и строительным материалом, служащим для ее реализации, то есть для конструирования пептидов – аминокислотами.
Роль транспортных РНК в синтезе белка была постулирована еще до их открытия. В 1955г. Крик приписал им функцию адаптера, который может нести аминокислоту и образовывать водородные связи с кодирующей полинуклеотидной матрицей. Гипотеза адаптера оказалась необходимой в связи с упоминавшейся уже невозможностью обнаружить между аминокислотами и нуклеиновыми кислотами стереохимическое соответствие, достаточное для того, чтобы обеспечить считывание генетического кода . В 1957г. в лаборатории Мэлона Хогланда было показано, что в ходе белкового синтеза активированные аминокислоты переносятся на особый тип РНК, получивший тогда наименование растворимой РНК и называемой теперь транспортной.
Стереохимия тРНК хорошо изучена и весьма характерна. Мы остановимся на ней поподробнее. Типичная молекула тРНК – это полинуклеотидная цепь длиной 75—90 (по преимуществу, 76) нуклеотидов. Молекулярные массы тРНК лежат в пределах 17.000—35.000. Часть оснований нуклеотидных пар, уже после синтеза тРНК, в определенных положениях модифицирована, это неканонические, редкие, так называемые ( минорные ), составляющие до 10% от общего числа. Среди них – дигидроуридин ( D ), псевдоуридин ( Ψ ) и инозин ( I ); последний играет существенную роль в узнавании кодона. В дополнение к этим модификациям несколько нуклеозидов метилированы. Все эти модификации – результат посттранскрипционного процессинга тРНК, которая копируется с «нормальной» матрицы. В 75% случаев молекулы тРНК открываются5′-гуанином (он фосфорилирован) и во всех случаях завершается триплетом ССА -3′.
Вторичная структура этой молекулы сформирована четырьмя короткими двуцепочечными стеблями и напоминает клеверный лист.
Каждый из четырех стеблей состоит из 4—7 уотсон-криковских пар, образующих двойные спирали. Сами стебли носят названия акцепторного, антикодонного, а также D (содержащий дигидроуридин) и T (содержащий риботимидин). Некоторые нуклеотиды консервативны, и их позиции в составе тРНК остаются инвариантными – либо полуинвариантными, если сохраняется их пуриновая или пиримидиновая природа. На акцепторном стебле тРНК имеется участок связывания с аминокислотой; он неспецифичен и для всех аминокислот один и тот же: ССА -3». Противоположный стебель содержит одноцепочечную петлю с антикодоном, распознающим кодон на мРНК. Две другие, боковые, петли предназначены для связывания с рибосомой и с аминоацил-тРНКсинтетазой (АРСазой). Четвертая, не всегда выраженная, петля так и называется – дополнительная или вариабельная (V). У тРНК, узнаваемых АРСазами класса I, она, как правило, короче (4—5 нуклеотидов), у тРНК, узнаваемых АРСазами класса II – длиннее (13—21 нуклеотидов).
Третичная (пространственная) структура любой тРНК складывает все ее четыре ветви (стебли с петлями) в так называемую Г-форму (L-форму, если использовать латиницу):
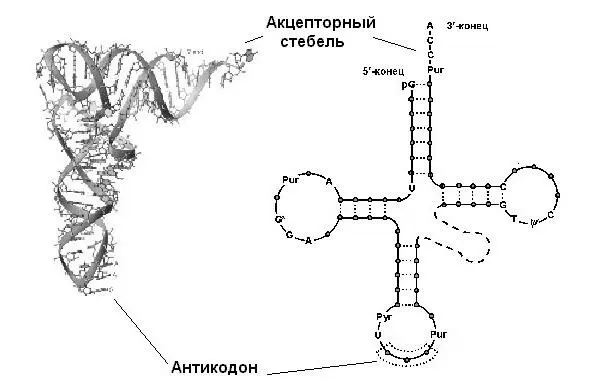
Г-форма состоит из двух почти перпендикулярных друг другу спиралей А-РНК (11 пар оснований на виток). Два отрезка буквы Г образованы ССА -3`-концом и антикодонной петлей, которые находятся на расстоянии 80Å друг от друга. Наружный край угла буквы Г образован Т-петлей. Акцепторный и Т-стебли уложены один вслед за другим и образуют единую двойную спираль. В примерно такую же структуру (только с расхождением осей на 26°) уложены антикодонный и D-стебли. Эта структура на предыдущем рисунке обозначена жирной черной кривой.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Клеймо создателя»
Представляем Вашему вниманию похожие книги на «Клеймо создателя» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.









![Наталья Александрова - Клеймо сатаны [= Табакерка Робеспьера] [litres]](/books/404375/natalya-aleksandrova-klejmo-satany-tabakerka-ro-thumb.webp)

Обсуждение, отзывы о книге «Клеймо создателя» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.