Сергей Ястребов - От атомов к древу. Введение в современную науку о жизни
Здесь есть возможность читать онлайн «Сергей Ястребов - От атомов к древу. Введение в современную науку о жизни» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 2018, ISBN: 2018, Издательство: Альпина нон-фикшн, Жанр: Биология, sci_biochem, sci_popular, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:От атомов к древу. Введение в современную науку о жизни
- Автор:
- Издательство:Альпина нон-фикшн
- Жанр:
- Год:2018
- Город:Москва
- ISBN:978-5-9614-5286-0
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
От атомов к древу. Введение в современную науку о жизни: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «От атомов к древу. Введение в современную науку о жизни»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Рекомендуется широкому кругу читателей, всерьез интересующихся современной биологией.
От атомов к древу. Введение в современную науку о жизни — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «От атомов к древу. Введение в современную науку о жизни», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Тем не менее считать, что модель Кунина опровергнута, пока не стоит. Ведь если архейная клетка к моменту захвата митохондрий уже имела множество бактериальных генов, то возникает вопрос: откуда она этих генов набралась? А как раз модель Кунина дает на этот вопрос очень внятный ответ. Кунин считает, что предком эукариот была неподвижная архея, имевшая зачатки цитоскелета и в связи с этим создавшая крупную «ветвящуюся» клетку со множеством складок внешней поверхности. В этих-то складках и поселились многочисленные бактерии — как альфа-протеобактерии, так и разные другие. Альфа-протеобактерии в дальнейшем были поглощены архейной клеткой и стали митохондриями — вот это, скорее всего, и запустило новый этап миграции их генов в ядро. Все остальные бактерии не были заключены внутри архейной клетки и эндосимбионтами не стали (скорее всего, потому, что архейной клетке это было не столь выгодно), но своими генами все равно успели щедро поделиться.
При всем этом не исключен (во всяком случае, не может считаться опровергнутым) и такой сценарий, согласно которому альфа-протеобактериальных симбионтов было два — древний и более поздний, причем митохондрии произошли от последнего. Эту гипотезу мы подробно обсудим в главе 14.
Предок, вывернутый наизнанку
Дальнейшим развитием «кунинского сценария» стала еще более парадоксальная гипотеза, согласно которой основная часть предковой архейной клетки соответствует ядру, и только ему. Цитоплазма же, по этой гипотезе, произошла от слившихся между собой внешних выростов архейной клетки, которые поначалу служили для обмена веществами с наружными симбионтами — предками митохондрий (см. рис. 10.9). Эту гипотезу предложили в 2014 году два американских биолога, двоюродные братья Дэвид и Базз Баумы [80] Baum D., Baum B. An inside-out origin for the eukaryotic cell // BMC Biology , 2014, V. 12, № 1, 76.
[81] Baum D., Baum B. The world in a cell // New Scientist , 2015, V. 225, № 3008, 28–29.
.
Гипотеза Баумов сразу объясняет, почему от архей у эукариот остались в основном белки, связанные с генетическими процессами. Да потому, что остаток собственно архейной клетки — это ядро! Кроме того, из новой гипотезы следует, что эндоплазматическая сеть — это на самом деле участок внешнего пространства, охваченный сомкнувшимися разрастаниями первичной клетки. Тогда получается, что внутри эндоплазматической сети могут найтись остатки архейной клеточной стенки. И действительно, там обнаружены белки, очень похожие на такие остатки. Это так называемые N-гликозилированные белки, в которых к боковым цепям аминокислоты аспарагина через атом азота присоединены остатки сахаров [82] Albers S. V., Meyer B. H. The archaeal cell envelope // Nature Reviews. Microbiology , 2011, V. 9, 414–426.
. То, что эти белки найдены у эукариот не на поверхности клетки, а глубоко в полости ЭПС, в самом деле заставляет очень серьезно задуматься.
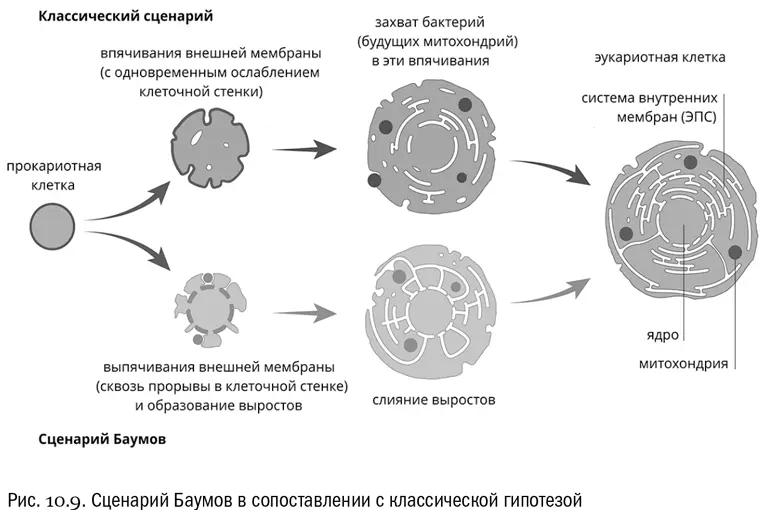
Ну а происхождение митохондрий в новой версии уж точно выглядит совершенно не связанным с фагоцитозом. Хлоропласты — иное дело, но они и приобретены были намного позже. Впрочем, в любом случае надо иметь в виду, что если само по себе симбиотическое происхождение митохондрий — твердо установленный факт, то все подробности того, как это случилось, относятся к области гипотез, которые пока что проверяются [83] Хороший обзор гипотезы Баумов на русском языке: https://postnauka.ru/faq/35994
. В главе 15 мы еще вернемся к этой теме.
Дела генетические
Между прокариотами и эукариотами есть несколько серьезных отличий, касающихся не столько устройства клетки (которое можно воочию увидеть под микроскопом), сколько принципов функционирования генетического аппарата. Перечислим важнейшие из этих отличий очень кратко.
Во-первых, прокариоты и эукариоты различаются способом «упаковки» генов в геном. Молекулу ДНК, заключающую в себе весь геном либо его существенную часть и связанную тем или иным способом со специальными белками, принято называть хромосомой . У прокариот хромосома чаще всего одна, а у эукариот почти всегда несколько, и они линейные, а не кольцевые.
Во-вторых, у эукариот полностью разобщены в пространстве процессы транскрипции и трансляции. При наличии ядра иначе и быть не может. У прокариот вполне возможна ситуация, когда на торчащий «хвост» информационной РНК, синтез которой еще продолжается, сразу же садится рибосома и начинает трансляцию. У эукариот такое полностью исключено. Транскрипция идет только в ядре, трансляция — только в цитоплазме. Даже те белки, которые в силу своих функций используются исключительно внутри ядра (например, гистоны), у эукариот синтезируются в цитоплазме и потом переправляются в ядро через ядерные поры.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «От атомов к древу. Введение в современную науку о жизни»
Представляем Вашему вниманию похожие книги на «От атомов к древу. Введение в современную науку о жизни» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.










Обсуждение, отзывы о книге «От атомов к древу. Введение в современную науку о жизни» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.