Сергей Ястребов - От атомов к древу. Введение в современную науку о жизни
Здесь есть возможность читать онлайн «Сергей Ястребов - От атомов к древу. Введение в современную науку о жизни» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 2018, ISBN: 2018, Издательство: Альпина нон-фикшн, Жанр: Биология, sci_biochem, sci_popular, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:От атомов к древу. Введение в современную науку о жизни
- Автор:
- Издательство:Альпина нон-фикшн
- Жанр:
- Год:2018
- Город:Москва
- ISBN:978-5-9614-5286-0
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
От атомов к древу. Введение в современную науку о жизни: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «От атомов к древу. Введение в современную науку о жизни»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Рекомендуется широкому кругу читателей, всерьез интересующихся современной биологией.
От атомов к древу. Введение в современную науку о жизни — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «От атомов к древу. Введение в современную науку о жизни», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Новый сценарий предполагает примерно следующую последовательность событий [77] Yutin N. et al. The origins of phagocytosis and eukaryogenesis // Biology Direct , 2009, V. 4, № 1, 9.
:
1. Предком эукариот была древняя архея, лишенная клеточной стенки, зато имевшая зачатки актинового цитоскелета. Это позволило архейной клетке стать крупной и приобрести сложную форму.
2. В складках наружной мембраны крупной археи стали поселяться разнообразные бактерии, которые паразитировали на ней или просто находили удобное убежище.
3. Близость этих бактерий создала поток чужеродных генов, от которого архее надо было защищаться, чтобы ее собственный геном не разладился. Именно защищаясь от чрезмерного проникновения генетического материала бактерий-паразитов, архея выработала сложную систему внутренних мембран, усовершенствовала внутриклеточный транспорт и цитоскелет. А паразитизм в конце концов превратился в симбиоз.
4. Один из бактериальных сожителей оказался настолько полезным в плане получения энергии, что архея навсегда замкнула его внутри собственной клетки, превратив из внешнего симбионта во внутреннего. Так появились митохондрии.
5. Фагоцитоз возник только в самом конце этих событий, как побочный эффект от усложнения цитоскелета и появления системы рециклизации мембран (последнее совершенно необходимо при постоянном обороте временных мембранных пузырьков, с которым связана работа ЭПС и аппарата Гольджи).
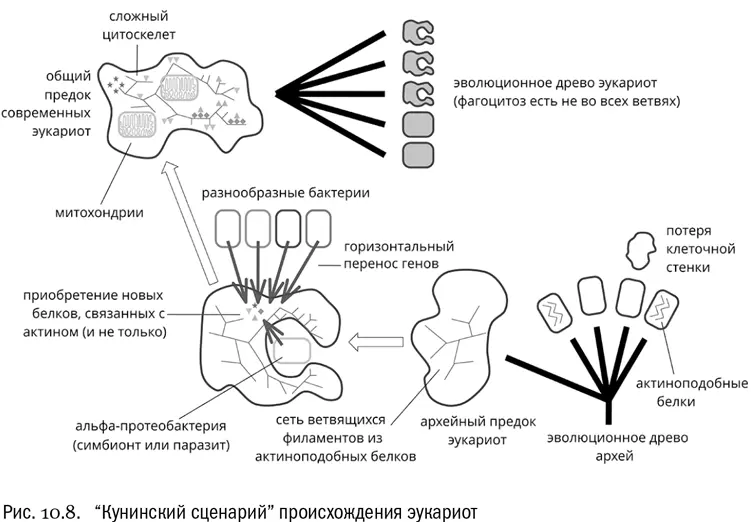
«Кунинский сценарий» имеет несколько красивых подтверждений. Например, известны современные морские археи с гигантскими клетками сложной формы, которые действительно покрыты снаружи симбиотическими бактериями. Причем эти внешние симбионты — не кто-нибудь, а протеобактерии, дальние родственники тех самых пурпурных бактерий, от которых произошли митохондрии [78] Muller F. et al. First description of giant Archaea (Thaumarchaeota) associated with putative bacterial ectosymbionts in a sulfidic marine habitat // Environmental Microbiology , 2010, V. 12, № 8, 2371–2383.
. Кроме того, сравнительный анализ белковых последовательностей показывает, что актиновый цитоскелет, скорее всего, сначала был неподвижным. Белки, позволяющие актиновым микрофиламентам еще и сокращаться, появились заметно позже. А это означает, что начаться прямо с фагоцитоза эволюция эукариот не могла.
Споры вокруг митохондрий
На самом деле сейчас конкурируют по меньшей мере два сценария происхождения эукариот: «раннемитохондриальный» (mito-early) и «позднемитохондриальный» (mito-late). Первый предполагает, что появление митохондрий было очень ранним событием, может быть даже запустившим эукариогенез как таковой. Второй гораздо ближе к уже рассмотренным классическим представлениям: сначала сформировалась полноценная эукариотная клетка с ядром, прочими мембранными структурами и фагоцитозом, а потом она проглотила будущую митохондрию и сделала ее своим симбионтом.
В последние несколько лет «раннемитохондриальная» гипотеза успела если не утвердиться, то, во всяком случае, набрать большую популярность. И, как и следовало ожидать, ее сразу начали подвергать скептическим проверкам. Например, недавняя работа испанских биоинформатиков показала, что эукариотные белки делятся по уровню древности на три группы [79] Pittis A. A., Gabaldon T. Late acquisition of mitochondria by a host with chimaeric prokaryotic ancestry // Nature , 2016, V. 531, 101–104.
.
К самой древней группе относятся белки архейного происхождения, в основном ядерные и (или) выполняющие функции, связанные с передачей генетической информации. Эти белки унаследованы прямо от архейного предка основной части эукариотной клетки, включающей ядро и цитоплазму. (Иногда эту часть клетки называют «ядерно-цитоплазматический компонент», ЯЦК.)
Более молодые белки имеют бактериальное происхождение и локализуются в цитоплазме, но не в ядре. Их функции часто связаны с мембранными структурами — эндоплазматической сетью (ЭПС) и аппаратом Гольджи. Причем вновь показано, что эти белки были заимствованы не от одной группы бактерий, а от нескольких разных — вероятнее всего, путем горизонтального переноса генов, когда архейный предок жил в составе какой-то сложной многовидовой колонии. Пока что, как видим, все соответствует модели Кунина.
А вот третья группа белков — самая молодая — происходит от альфа-протеобактерий и локализуется в митохондриях. Несомненно, они были приобретены вместе с бактериальными симбионтами, которые этими самыми митохондриями стали. Вот это уже противоречит модели Кунина, которая, совсем наоборот, предполагает, что симбиоз с предками митохондрий был «спусковым крючком эукариотизации»: сначала появились они, а потом уже ЭПС, ядро и все остальное. А тут получается, что субъектом, который приобрел митохондрии, была достаточно сложная клетка, уже являвшаяся генетической химерой, то есть совмещавшая в себе архейные и бактериальные гены.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «От атомов к древу. Введение в современную науку о жизни»
Представляем Вашему вниманию похожие книги на «От атомов к древу. Введение в современную науку о жизни» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.










Обсуждение, отзывы о книге «От атомов к древу. Введение в современную науку о жизни» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.