Сергей Ястребов - От атомов к древу. Введение в современную науку о жизни
Здесь есть возможность читать онлайн «Сергей Ястребов - От атомов к древу. Введение в современную науку о жизни» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 2018, ISBN: 2018, Издательство: Альпина нон-фикшн, Жанр: Биология, sci_biochem, sci_popular, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:От атомов к древу. Введение в современную науку о жизни
- Автор:
- Издательство:Альпина нон-фикшн
- Жанр:
- Год:2018
- Город:Москва
- ISBN:978-5-9614-5286-0
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
От атомов к древу. Введение в современную науку о жизни: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «От атомов к древу. Введение в современную науку о жизни»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Рекомендуется широкому кругу читателей, всерьез интересующихся современной биологией.
От атомов к древу. Введение в современную науку о жизни — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «От атомов к древу. Введение в современную науку о жизни», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Скорее всего, изменения в генетическом коде митохондрий стали накапливаться уже после того, как митохондрии окончательно стали внутриклеточными симбионтами. Со временем большая часть митохондриальных генов перешла в ядро, а оставшийся митохондриальный геном стал таким маленьким, что сбой его работы (который неизбежен при любом изменении генетического кода) перестал быть непременно смертоносным. Надо учитывать, что митохондриальные гены, мигрировавшие в ядро и вошедшие в состав ядерного генома, экспрессируются там, где они теперь находятся, то есть в ядре. А их белковые продукты (даже если эти белки нужны только для митохондрий) синтезируются на обычных эукариотных рибосомах, находящихся в цитоплазме, и внутрь митохондрий попадают уже оттуда, с помощью специальных переносчиков.
После того как в митохондриальном генетическом коде накопилось некоторое количество изменений, митохондриальные белки стало уже невозможно правильно синтезировать на эукариотных рибосомах. Вот тогда-то миграция митохондриальных генов в ядро и прекратилась. Если бы не это, митохондрии, вероятно, в конце концов совсем потеряли бы свой генетический аппарат, и тогда разгадать их симбиотическое происхождение было бы гораздо труднее.
И наконец, еще один важнейший факт состоит в том, что — как мы сейчас знаем — эукариоты эволюционно близки не к бактериям, а к археям. Это выяснилось еще тогда, когда археи как отдельная эволюционная ветвь были только открыты (см. главу 5). Скорее всего, эукариоты прямо от них и произошли. А вот предки митохондрий и хлоропластов, наоборот, не археи, а бактерии. Тут произошло довольно редкое событие, а именно слияние совершенно разных эволюционных ветвей.
«Первичный фагоцит»
Факты, которые мы теперь знаем, легко встраиваются в следующий довольно простой эволюционный сценарий (см. рис. 10.6). Когда-то в далекой древности (вероятно, два с лишним миллиарда лет назад) некая бесцветная архея приобрела сложный цитоскелет, включающий актино-миозиновый комплекс. Клеточную стенку она, наоборот, потеряла. Получился «первичный фагоцит» — подвижная клетка, способная активно ползать, выпуская ложноножки, и фагоцитировать (то есть заглатывать целиком) любые более мелкие объекты, в том числе и клетки бактерий. Это оказалось очень выгодным способом питания. Но такое питание непременно предполагает, что хищник должен быть крупнее жертвы. И действительно, современные эукариотные одноклеточные хищники — амебы и жгутиконосцы — по линейным размерам примерно в десять раз превосходят бактерии, которыми они питаются.
Оговорка про линейные размеры тут очень важна. Если увеличить линейный размер клетки в десять раз (обычный порядок разницы между эукариотами и прокариотами), то — по законам геометрии — ее объем увеличится примерно в 1000 раз, с пропорциональным ростом нагрузки на все внутриклеточные системы синтеза и транспорта. Вот для того, чтобы структурировать огромный внутренний объем, у предков эукариот и образовалась система внутренних мембран, включающая ЭПС, аппарат Гольджи, множество вакуолей и ядро. Возникший в результате организм представлял собой что-то вроде крупной бесцветной хищной амебы. Ничего особенного, подобные эукариоты есть и сейчас, разве что они нередко обладают еще и жгутиками в придачу к ложноножкам.
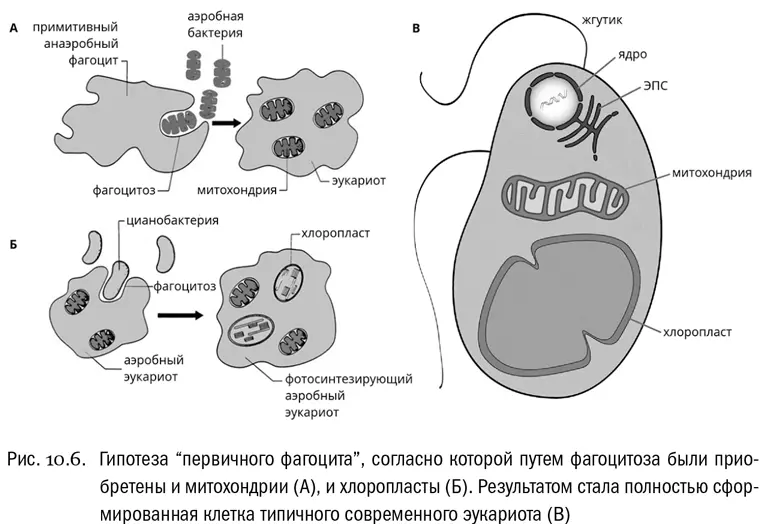
Питание путем фагоцитоза предопределило возможность появления у эукариот внутриклеточных сожителей, то есть, по-научному говоря, симбионтов. Для прокариот, лишенных способности заглатывать кого бы то ни было, приобретение симбионтов таким способом невозможно. А вот амебоидный эукариот может легко превратить некоторые захваченные клетки из пищи в симбионтов, если «примет решение» не переваривать их, а предоставить им жить в хозяйской клетке, делясь с хозяином полезными продуктами обмена веществ. В современных научных статьях такой захват эукариотами чужих клеток часто буквально называют порабощением (enslavement). Именно путем такого «порабощения» эукариотная клетка и приобрела сначала митохондрии, а потом хлоропласты.
К сожалению, на самом деле все не так просто. Познание природы имеет свои закономерности: как правило, чем больше мы узнаем, тем больше возникает новых вопросов, причем таких, которые на предыдущем уровне знания просто не могли бы никому прийти в голову. И проблема происхождения эукариот может послужить отличной иллюстрацией того, что так бывает очень часто. Мы сейчас увидим, что есть как минимум два соображения, заново затуманивающих, казалось бы, предельно ясно описанную картинку.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «От атомов к древу. Введение в современную науку о жизни»
Представляем Вашему вниманию похожие книги на «От атомов к древу. Введение в современную науку о жизни» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.










Обсуждение, отзывы о книге «От атомов к древу. Введение в современную науку о жизни» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.