Александр Марков - Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества
Здесь есть возможность читать онлайн «Александр Марков - Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 2019, ISBN: 2019, Издательство: АСТ, Corpus, Жанр: Биология, sci_popular, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.
- Название:Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества
- Автор:
- Издательство:АСТ, Corpus
- Жанр:
- Год:2019
- Город:Москва
- ISBN:978-5-17-114115-8
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Правда, листья у трансгенных A. thaliana не стали такими же сложными, как у сердечника C. hirsuta . Вторичные листочки не сформировались, вместо них получились лопасти. По-видимому, чтобы лопасти стали полноценными вторичными листочками с собственными черешками, помимо RCO нужно что-то еще — какие-то дополнительные генетические особенности, имеющиеся у сердечника и отсутствующие у резуховидки. Интересно, что точно такие же лопастные листья, как у трансгенных A. thaliana , характерны для родственного вида A. lyrata , в геноме которого, как мы уже упоминали, сохранился ген RCO .
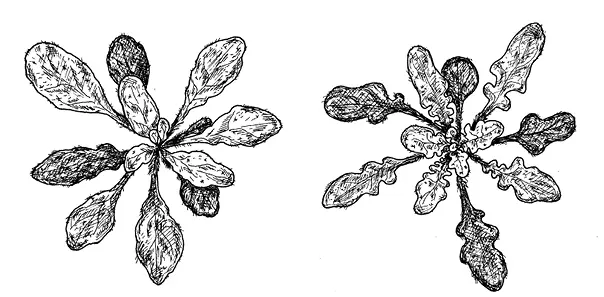
рис. 16.2.Ген RCO , вставленный в геном Arabidopsis thaliana , превращает простые листья в лопастные. Слева — обычное растение A. thaliana, справа — трансгенное. Длина масштабной линейки — 1 см.
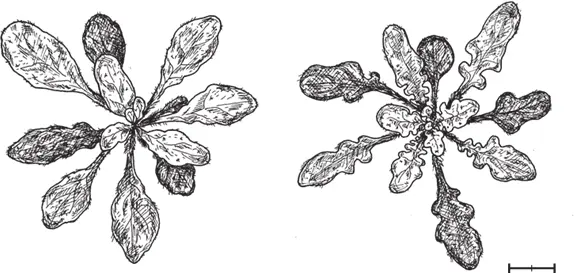
Дальнейшие опыты показали, что различные варианты гена LMI 1, взятые у разных представителей крестоцветных (и даже у некоторых других двудольных), вызывают формирование лопастных листьев у A. thaliana , если белок-кодирующие участки этих генов объединить с регуляторным участком RCO . Это еще один довод в пользу того, что ключевое эволюционное новшество при формировании сложных листьев у крестоцветных состояло в изменении регуляторного участка RCO , тогда как свойства белков, кодируемых этими генами, весьма консервативны.
Наконец, ученые попытались выяснить, каким образом ген RCO со своим специфическим регуляторным участком превращает простой лист с волнистым или зазубренным краем в лопастной или перистый. Для этого потребовалось внимательнее приглядеться к тому, что именно делает RCO , как он влияет на развитие листа. Исследователи сравнили рост листовых зачатков у обычного сердечника с перистыми листьями и у мутантов rco с простыми листьями. В обоих случаях на краю листовой пластинки формируются небольшие округлые выступы. Дальнейший рост этих выступов протекает одинаково у обычных C. hirsuta и у мутантов rco . Однако в поведении клеток, расположенных в промежутках между выступами, наблюдаются контрастные различия. В норме у C. hirsuta эти клетки не делятся. Поэтому между растущими выступами формируются глубокие вырезы, а сами выступы в итоге становятся вторичными листочками. У мутантов rco клетки в промежутках между выступами активно делятся, поэтому край листового зачатка растет равномерно. В результате вместо перистого листа получается сплошная листовая пластинка с волнистым краем. По-видимому, это значит, что ген RCO , экспрессирующийся в основаниях выступов, не столько ускоряет рост самих выступов, сколько подавляет деление клеток в промежутках между зонами своей экспрессии.
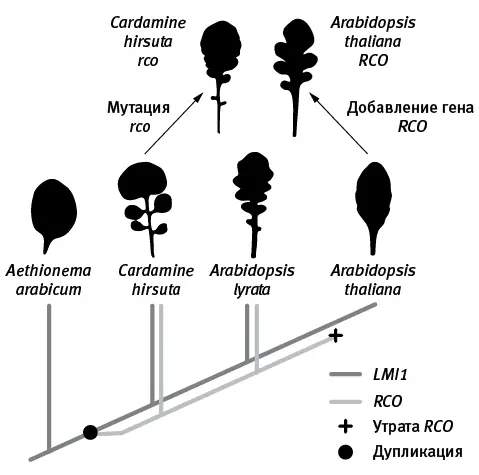
рис. 16.3.Эволюция формы листа у крестоцветных. Внизу — эволюционное дерево генов LMI1 ( темно-серые линии ) и RCO ( светло-серые ). У базальных крестоцветных ( Aethionema arabicum ) не было гена RCO , а листья были простыми. У общего предка Cardamine и Arabidopsis произошла дупликация гена LMI1 ( черный кружок ). Одна из двух копий удвоившегося гена сохранила старую функцию, а другая ( RCO ) приобрела новую благодаря изменениям регуляторного участка, что способствовало усложнению формы листьев. Впоследствии утрата RCO ( черный крестик ) привела к вторичному упрощению листьев у A. thaliana . Поломка гена RCO у C. hirsuta (мутация rco ) ведет к упрощению листьев. Возвращение утраченного гена RCO в геном A. thaliana приводит к усложнению формы листа. По рисунку из Vlad et al., 2014 .
Таким образом, детали головоломки постепенно сложились в целостную картину (она схематично изображена на рис. 16.3). У первых крестоцветных, унаследовавших ген LMI 1 от своих предков и еще не имевших RCO , листья были простыми, как у крылотычинника ( Aethionema ). Затем произошла дупликация LMI 1 с последующей неофункционализацией одной из копий. Новая функция возникла благодаря изменению регуляторной части гена, что повлияло на область его экспрессии. Ген начал экспрессироваться в основаниях выступов развивающейся листовой пластинки. Это затормозило деление клеток в промежутках между выступами и привело к усложнению формы листа. В дальнейшем предки A. thaliana утратили ген RCO , что вызвало вторичное упрощение листа (как у C. hirsuta с мутацией rco ). Однако в геноме A. thaliana сохранились все прочие необходимые компоненты программы развития сложного листа, не хватает лишь гена RCO (или любого похожего гена, управляемого регуляторной областью RCO ). Если его искусственно вставить в геном A. thaliana , растение приобретает сложные лопастные листья, в точности как у родственного вида A. lyrata .
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества»
Представляем Вашему вниманию похожие книги на «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.










Обсуждение, отзывы о книге «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.