Александр Марков - Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества
Здесь есть возможность читать онлайн «Александр Марков - Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 2019, ISBN: 2019, Издательство: АСТ, Corpus, Жанр: Биология, sci_popular, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.
- Название:Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества
- Автор:
- Издательство:АСТ, Corpus
- Жанр:
- Год:2019
- Город:Москва
- ISBN:978-5-17-114115-8
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
К числу эукариотических инноваций, наиболее трудных для объяснения, относится эукариотический секс (называемый также амфимиксисом, или попросту половым размножением). Это специфический и очень эффективный способ перемешивания генетического материала разных особей, включающий сингамию (слияние гаплоидных клеток или ядер, приводящее к удвоению хромосомного набора) и мейоз (особый вариант клеточного деления, приводящий к сокращению числа хромосом вдвое). Мейоз сопровождается кроссинговером , в ходе которого гомологичные хромосомы обмениваются гомологичными участками.
Прокариотический секс (горизонтальный перенос генов, ГПГ) устроен гораздо проще. Можно назвать три основных отличия эукариотического секса (полового размножения) от прокариотического (ГПГ).
Во-первых, у эукариот обмен генами взаимный, двусторонний: каждый участник является и донором, и реципиентом. У прокариот передача генетического материала строго однонаправленная: один участник — донор ДНК, другой — реципиент.
Во-вторых, для эукариот характерна полногеномная рекомбинация: в обмене участвуют два полных генома. У прокариот в рекомбинации участвует один полный геном и один фрагмент.
В-третьих, у эукариот все гены, участвующие в рекомбинации, сохраняют шанс перейти к потомкам. У прокариот фрагмент ДНК донора либо встраивается в геном реципиента вдобавок к уже имеющимся там генам (но этот вариант не может использоваться регулярно, например в каждом поколении, ведь это привело бы к безудержному разрастанию генома), либо замещает собой гомологичный фрагмент генома реципиента. При этом замещенные аллели реципиента уничтожаются и теряют шанс перейти к потомкам (рис. 5.1).
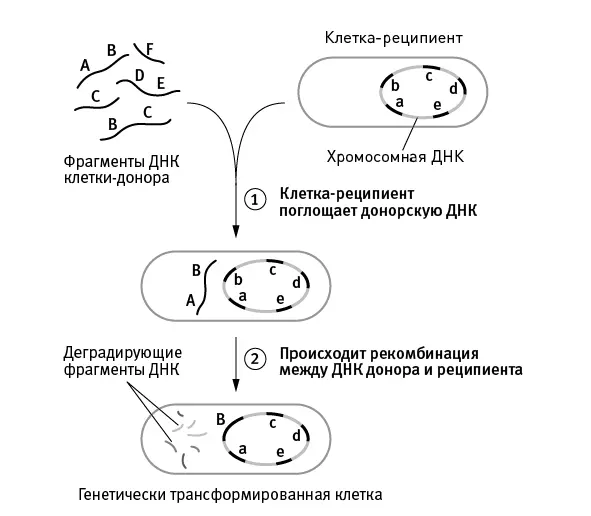
рис. 5.1.Один из способов горизонтального переноса генов у прокариот (естественная трансформация, сопряженная с гомологичной рекомбинацией), наиболее близкий по своим эволюционным последствиям к эукариотическому сексу. Бактерия поглощает фрагмент чужой ДНК из окружающей среды (это могут быть, например, фрагменты геномов погибших бактерий того же вида), а затем встраивает его в свою кольцевую хромосому вместо собственного гомологичного фрагмента. В результате аллели бактерии-реципиента уничтожаются, замещаясь чужими аллелями. В данном случае аллель B встроился в хромосому и будет передан потомкам, а аллель b погиб.
Раньше многие специалисты предполагали, что первые эукариоты были бесполыми (размножались клонально — простым делением — и не имели полового процесса), а потом у кого-то из них появилось половое размножение. В таком случае проблема возникновения эукариотического секса не связана напрямую с проблемой происхождения эукариот. Однако по мере накопления геномных данных стало ясно, что эукариотический секс является одной из фундаментальных особенностей эукариот, которая почти наверняка имелась уже у LECA. В частности, появляется все больше аргументов в пользу того, что все современные бесполые группы эукариот произошли от предков, имевших мейоз (в их геномах сохранились остатки необходимых для мейоза генов), а значит, и половой процесс. Стало быть, происхождение эукариот и происхождение полового размножения — проблемы взаимосвязанные.
Половой процесс солелюбивой археи Haloferax .Если эукариотический секс — исходный признак эукариот, то имеет смысл поискать среди современных архей варианты генетического обмена, переходные между типичным прокариотическим и эукариотическим сексом. Крайне интересно было бы выяснить, как меняются генами асгардархеи (переходная группа между обычными археями и эукариотами), но об этом пока ничего не известно. На сегодняшний день подробно изучена система генетического обмена только у одной археи — солелюбивого (галофильного) микроба Haloferax volcanii , обитателя Мертвого моря. У Haloferax обнаружена система передачи генетического материала, действительно похожая на что-то промежуточное между обычным прокариотическим ГПГ (например, конъюгацией бактерий) и эукариотическим сексом.
Клетки Haloferax спариваются, соединяясь цитоплазматическими мостиками, по которым геномная ДНК может передаваться в обе стороны, то есть обе клетки могут быть как донорами, так и реципиентами генетического материала. При этом могут передаваться и крупные фрагменты хромосомы (длиной до 13 % генома как минимум). В лаборатории можно добиться даже слияния клеток: для этого понижают концентрацию ионов Mg 2+ в среде, что приводит к растворению клеточных стенок и дестабилизации цитоплазматических мостиков. Предполагается, что такое слияние иногда происходит и в природе.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества»
Представляем Вашему вниманию похожие книги на «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.










Обсуждение, отзывы о книге «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.