Александр Марков - Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества
Здесь есть возможность читать онлайн «Александр Марков - Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 2019, ISBN: 2019, Издательство: АСТ, Corpus, Жанр: Биология, sci_popular, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.
- Название:Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества
- Автор:
- Издательство:АСТ, Corpus
- Жанр:
- Год:2019
- Город:Москва
- ISBN:978-5-17-114115-8
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
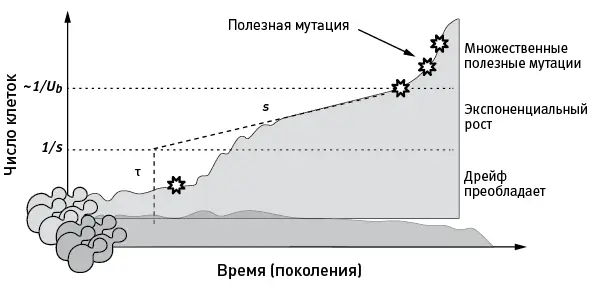
рис. 4.1.Типичная судьба клонов бесполых организмов в ходе адаптации. Темно-серым цветом показана динамика численности клона дрожжей, в котором не возникло полезных мутаций. Такой «нейтральный» клон, скорее всего, не достигнет высокой численности и будет вытеснен более успешными конкурентами. Светло-серым цветом показана судьба клона, в котором в момент времени τ возникла полезная мутация (изображена звездочкой ). Пока численность такого клона мала, он может вымереть просто случайно (из-за дрейфа). Чтобы отбор начал поддерживать носителей полезной мутации, клон должен достичь (опять-таки случайно) пороговой численности, которая для мутации с полезным эффектом s примерно равна 1/ s . После прохождения этого порога клон с полезной мутацией вступает в фазу экспоненциального роста численности. Когда численность клона приблизится к 1/ U b , где U b — частота возникновения полезных мутаций, в нем с большой вероятностью начнут появляться дополнительные полезные мутации. Шкала по вертикальной оси логарифмическая. По рисунку из Levy et al., 2015 .
После того как был получен гигантский массив данных по скоростям роста численности отдельных клонов (а мы помним, что их полмиллиона в каждой из двух подопытных популяций), в дело пошла статистика и компьютерное моделирование. Выводы получились следующие.
Численность большинства клонов с течением времени снижалась, нередко до полного исчезновения клона. Такая судьба характерна для «нейтральных» клонов, в которых не возникло полезных мутаций. Однако примерно у 5 % клонов за первую сотню поколений появилась и «установилась» хотя бы одна полезная мутация.
По мере размножения клеток с полезными мутациями средняя приспособленность популяции росла. Это, в свою очередь, приводило к тому, что «нейтральные» клоны, в которых не было полезных мутаций, оказывались во все более проигрышном положении. Их численность продолжала убывать. Более того, многие «удачливые» клоны, поначалу бодро размножавшиеся благодаря рано возникшей слабополезной мутации, в дальнейшем приходили в упадок и вымирали. Это объясняется тем, что на фоне возросшей приспособленности конкурентов их полезная мутация уже не давала преимущества.
Средняя приспособленность популяций за 100 поколений выросла на 5 %. Соответственно, к этому времени все обладатели мутаций с s < 0,05 оказались в проигрыше, хотя на ранних этапах именно эти многочисленные клоны вносили основной вклад в общий рост приспособленности.
Эти результаты говорят о том, что ключевым процессом, определяющим эволюционную динамику большой бесполой популяции, является безжалостная конкуренция между клонами — обладателями разных полезных мутаций (клональная интерференция). При этом ранними стадиями адаптации заправляют высоковероятные и потому многочисленные слабополезные мутации. Таких мутаций, повышающих приспособленность на 2–5 % (0,02 < s < 0,05, мутации с более слабым эффектом остались незамеченными), в первой из двух подопытных популяций установилось около 20 000, во второй — 11 000. На этом этапе ход адаптации в значительной мере предсказуем: его можно описать уравнениями и при повторных эволюционных экспериментах результаты получаются схожими. Затем, по мере роста средней приспособленности, многочисленные слабополезные мутации фактически перестают быть полезными и начинают отбраковываться. Продолжает нарастать лишь численность тех немногих клонов, в которых возникли редкие мутации с сильным положительным эффектом. К 70-му поколению динамика общей приспособленности определялась в основном мутациями с полезностью около 0,1. Таких мутаций в двух подопытных популяциях было 5000 и 3000 соответственно. После 110-го поколения на первый план вышли примерно по 100 клонов с наиболее полезными мутациями ( s > 0,1). На этом этапе ход адаптации становится все менее предсказуемым, поскольку то, когда и в каком количестве возникнут эти маловероятные мутации, зависит от случая.
Внимательный читатель может заметить кажущееся противоречие между этими результатами и теми, которые были получены в эксперименте Ричарда Ленски (см. Исследование № 1). Там сначала закреплялись мутации с сильным положительным эффектом, а лишь затем начинали фиксироваться слабополезные. На самом деле расхождений тут нет, просто речь идет о разном временном масштабе и о разных этапах адаптации: у Ленски — десятки тысяч поколений, в рассматриваемой работе — 168. Иными словами, в данном исследовании внимание фокусируется на том начальном этапе, который Ленски пролетел на полном ходу. Разнятся также отслеживаемые параметры и разрешающая способность методов. В работе Ленски регистрировались уже зафиксировавшиеся, то есть достигшие стопроцентной частоты, полезные мутации, а здесь говорится о мутациях, только начавших свой путь. Подавляющее большинство из них никогда не зафиксируется, а будет вытеснено в ходе клональной интерференции. Если бы эксперимент продлился еще несколько тысяч поколений, то зафиксировались бы как раз немногочисленные мутации с самым сильным положительным эффектом, точно как в эксперименте Ленски.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества»
Представляем Вашему вниманию похожие книги на «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.










Обсуждение, отзывы о книге «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.