Владимир Бердников - Эволюция и прогресс
Здесь есть возможность читать онлайн «Владимир Бердников - Эволюция и прогресс» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Новосибирск, Год выпуска: 1991, ISBN: 1991, Издательство: Наука, Жанр: Биология, sci_popular, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Эволюция и прогресс
- Автор:
- Издательство:Наука
- Жанр:
- Год:1991
- Город:Новосибирск
- ISBN:5-02-030062-4
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Эволюция и прогресс: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Эволюция и прогресс»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Книга предназначена для биологов широкого профиля, а также всех интересующихся вопросами эволюции живых существ.
Эволюция и прогресс — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Эволюция и прогресс», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Основные функции многоклеточного организма в значительной мере обеспечиваются его надмолекулярными структурами. Каждая из них возникает в ходе индивидуального развития под контролем своих специальных генов, работа которых так тонко увязана между собой (в программу развития), что изменение экспрессии любого из них немедленно отражается на функции взрослой структуры. Фактически это означает, что возникающие с повышенной частотой мутации по генной экспрессии начинают выступать в роли мутаций, ослабляющих или усиливающих мощность рабочих структур. Отметим, что изменения наследственного материала, ведущие к увеличению числа специальных генов надклеточных рабочих структур, можно с полным основанием отнести к мутациям, повышающим «выход» благоприятных мутаций.
Математическая модель мобилизующего отбора
Попробуем более формально проанализировать механизм действия мобилизующего отбора. Представим себе, что в какой-то адаптивной зоне обитает большое число родственных видов, обладающих одним и тем же признаком — гомологичной и изофункциональной рабочей структурой. Пусть величина этого признака (под которым в данном случае мы понимаем логарифм мощности структуры) определяется суммарным вкладом его специальных генов. Каждый из них может находиться в двух аллельных состояниях 1 и 0. Аллель 1 соответствует активному состоянию гена (когда он экспрессируется), аллель 0 — отсутствию экспрессии. Таким образом, величина признака (Z) определяется числом специальных генов, находящихся в активном состоянии. Полное число генов (G) в геноме разных видов постоянно, тогда как число специальных генов (N ), определяющих степень развития признака, различно. Гены, не вошедшие в состав специальных (их число равно G — N ), будем называть «сторонними». Они определяют развитие каких-то других (сторонних) признаков.
В среде обитания время от времени происходят медленные однонаправленные изменения, когда над каждым видом нависает реальная угроза гибели. Избежать вымирания вид может за счет компенсаторного изменения величины признака на какое-то число единиц. (Что соответствует увеличению мощности рабочей структуры в какое-то число раз.) Увеличение признака на одну единицу может произойти за счет фиксации мутаций двух типов. Во-первых, это могут быть мутации, активирующие специальные гены, и, во-вторых, мутации, переводящие ген из разряда «сторонних» в разряд специальных (с аллельным состоянием 1 ). Точно так же сдвиг признака в сторону его уменьшения может быть достигнут либо инактивацией одного специального гена, либо переводом активного специального гена в состав «сторонних».
Для своего филетического выживания каждый вид должен за единицу макроэволюционного времени осуществить фиксацию нескольких благоприятных мутаций (любого типа), изменяющих признак на требуемую величину ΔZ. Донорами этих мутаций являются чаще всего специальные гены признака, так как изменение экспрессии гена — более вероятное событие, чем его перепрофилирование. Соответственно вероятность филетического выживания вида (его адаптируемость) будет расти с увеличением числа специальных генов (N ). Это создает предпосылки роста генетической сложности признака.
Такая простая модель позволяет осуществить машинный эксперимент. На каждом шаге модели в окружающей среде каждого вида происходят случайные изменения, которые никак не связаны с изменением среды обитания остальных видов. От одних видов среда требует увеличения признака, а от других — уменьшения. Всю эту картину можно передать типом распределения величины ΔZ. В данном эксперименте мы остановились на биномиальном симметричном распределении. С подробностями вычисления вероятностей выживания видов в новых условиях среды можно ознакомиться в книге «Основные факторы макроэволюции».
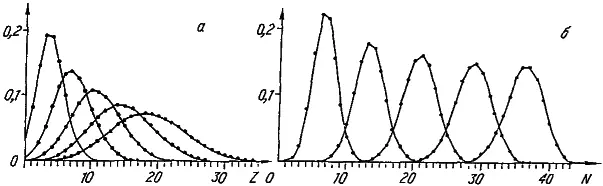
Рис. 27. Моделирование динамики видовых распределений по величине признака (а) и числу его специальных генов (б). Приведенные распределения (слева направо) соответствуют 20, 50, 100, 200, 800-му шагу модели. Последнее распределение финальное.
Проследим за изменением видовых распределений по величине признака (рис. 27, а ). В начальный момент все виды обладают минимальным значением признака, поскольку его развитие обеспечивает всего один специальный ген. Однако завершается машинная эволюция широким финальным распределением со средним значением, равным 18,4. Таким образом, несмотря на то, что среда с равной вероятностью благоприятствует как увеличению, так и уменьшению значения признака, его средневидовая величина () неуклонно растет. Правда, параллельно с этим нарастает и уровень межвидовых различий по степени развития признака (σ z) (рис. 28). В итоге даже в финальном распределении после долгой эволюции сохраняется довольно много видов с «архаичной», «примитивной», величиной признака.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Эволюция и прогресс»
Представляем Вашему вниманию похожие книги на «Эволюция и прогресс» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.









Обсуждение, отзывы о книге «Эволюция и прогресс» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.