Владимир Бердников - Эволюция и прогресс
Здесь есть возможность читать онлайн «Владимир Бердников - Эволюция и прогресс» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Новосибирск, Год выпуска: 1991, ISBN: 1991, Издательство: Наука, Жанр: Биология, sci_popular, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Эволюция и прогресс
- Автор:
- Издательство:Наука
- Жанр:
- Год:1991
- Город:Новосибирск
- ISBN:5-02-030062-4
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Эволюция и прогресс: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Эволюция и прогресс»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Книга предназначена для биологов широкого профиля, а также всех интересующихся вопросами эволюции живых существ.
Эволюция и прогресс — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Эволюция и прогресс», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Другой сильный аргумент против ортоселекции был представлен американским палеонтологом Дж. Цисне, проследившим за ростом дифференциации конечностей у водных членистоногих. Для количественной оценки этой формы морфофункционального прогресса Цисне предложил использовать степень тагмозиса конечностей (h), которую можно вычислить по формуле (2.1). Каждый отряд водных членистоногих (от примитивных трилобитов до высших ракообразных) был охарактеризован величиной h, после чего было проанализировано, как в течение 600 млн лет эволюции этих беспозвоночных изменялся средний для отряда уровень тагмозиса.
Оказалось, что в ходе исторического развития водных членистоногих величина возрастала, следуя S-образной (логистической) кривой (рис. 24). В течение первых 250 млн лет шло довольно быстрое увеличение среднего тагмозиса, затем этот процесс стал тормозиться и практически прекратился к началу мезозоя. Так что за последние 150 млн лет уровень не повышался, хотя одни отряды вымирали, а другие возникали.
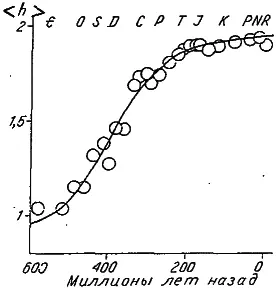
Рис. 24. Рост среднего тагмозиса () конечностей водных членистоногих в историческом развитии (по: [Cisne, 1974]).
E — кембрий, О — ордовик, S — силур, Р — палеоген, N — неоген, R — современность. Остальные условные обозначения см. рис. 22.
Данное явление нельзя объяснить исчерпанием возможностей к прогрессивной дифференциации конечностей, так как средний тагмозис на стадии плато ( = 1,8) намного ниже, чем у десятиногих ракообразных (h = 2,5). Низкий уровень плато объясняется существованием в течение многих миллионов лет практически постоянной доли «примитивных» отрядов. Это плато продолжительностью в 150 млн лет явно свидетельствует против таинственных внутренних (гомогенетических) факторов, толкавших членистоногих по пути прогресса, но, самое главное, оно ставит серьезные трудности и перед сторонниками ортоселекции. Ведь выгода от функциональной дифференциации конечностей, казалось бы, очевидна. Тогда почему же доля примитивных отрядов не убывала в течение последних 150 млн лет?
В 1949 г. Т. Уэстолл предпринял попытку измерить темп прогрессивной эволюции у двоякодышащих рыб. В данном случае уровень организации оценивался числом «прогрессивных» структурных новшеств (числом баллов). Динамика прогресса структурной сложности имеет тот же S- образный вид: период быстрого увеличения числа баллов продолжительностью около 150 млн лет сменяется гораздо более долгим плато (рис. 25).
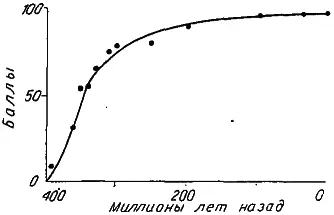
Рис. 25. Увеличение числа прогрессивных признаков (в баллах) в историческом развитии двоякодышащих рыб (по: [Рефф, Кофмен, 1986]).
Характер кривых, передающих три случая морфологического прогресса (рост тагмозиса у членистоногих, сложности лопастной линии у аммоноидей и продвинутости организации у двоякодышащих рыб) весьма сходен. Во всех случаях первоначальный период подъема сменяется периодом застоя. Объяснение этому удивительному феномену мы попытаемся дать в следующей главе.
Глава 6. Мобилизующий отбор
Эволюция подотряда Ammonitina протекала необычайно бурно. Все морфологические особенности раковины — и внешняя форма, и скульптура, и перегородки — проявляли при этом сильнейшую мобильность. <���…> Способы усложнения лопастной линии были также необычайно многообразны.
Б.Е. РуженцевМутабильность
К проблеме генетической сложности структуры можно подойти еще с одной стороны. Дело в том, что специальные гены, как и всякие другие, обязаны мутировать, порождая внутрипопуляционную изменчивость по мощности соответствующих рабочих структур. Заметим, что мощность морфологической структуры, с точки зрения генетика, следует отнести к мерным (количественным) признакам, для которых типичен непрерывный характер изменчивости. Мутации по этим признакам можно разделить на две четко очерченные категории — с качественным и количественным эффектом. Первые вызывают настолько резкое изменение значения признака, что наблюдатель видит их эффект без всяких измерений. Примерно также проявляет себя и потеря гена, поэтому в основе мутаций с качественным эффектом обычно лежит серьезное повреждение функции гена.
Мутации второй категории ведут к небольшому (на несколько процентов) сдвигу величины признака, и этот эффект, как правило, меньше шума среды. Еще одна особенность данных мутаций заключается в том, что они могут не только снижать, но и повышать значение признака. Поэтому природные популяции нередко хорошо отвечают на отбор по мерным признакам в обоих направлениях. За мутациями этого типа и за возникающими в результате их действия аллелями исторически закрепилось несколько неудачное название полигенных.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Эволюция и прогресс»
Представляем Вашему вниманию похожие книги на «Эволюция и прогресс» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.









Обсуждение, отзывы о книге «Эволюция и прогресс» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.