Charles J. Dorman - Structure and Function of the Bacterial Genome
Здесь есть возможность читать онлайн «Charles J. Dorman - Structure and Function of the Bacterial Genome» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.
- Название:Structure and Function of the Bacterial Genome
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Structure and Function of the Bacterial Genome: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Structure and Function of the Bacterial Genome»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Structure and Function of the Bacterial Genome Blends knowledge of gene regulatory mechanisms with a consideration of nucleoid structure and dynamics Offers a 'DNA-centric' approach to considering transcription control Views horizontal gene transfer from a gene regulation perspective Assesses the opportunities and limitations of designing synthetic microbes or rewiring existing ones
is an ideal book for graduate and undergraduate students studying microbial cell biology, bacterial pathogenesis, gene regulation, and molecular microbiology. It will also appeal to principal investigators conducting research on these and related topics and researchers in synthetic biology and other arms of biotechnology.
Structure and Function of the Bacterial Genome — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Structure and Function of the Bacterial Genome», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
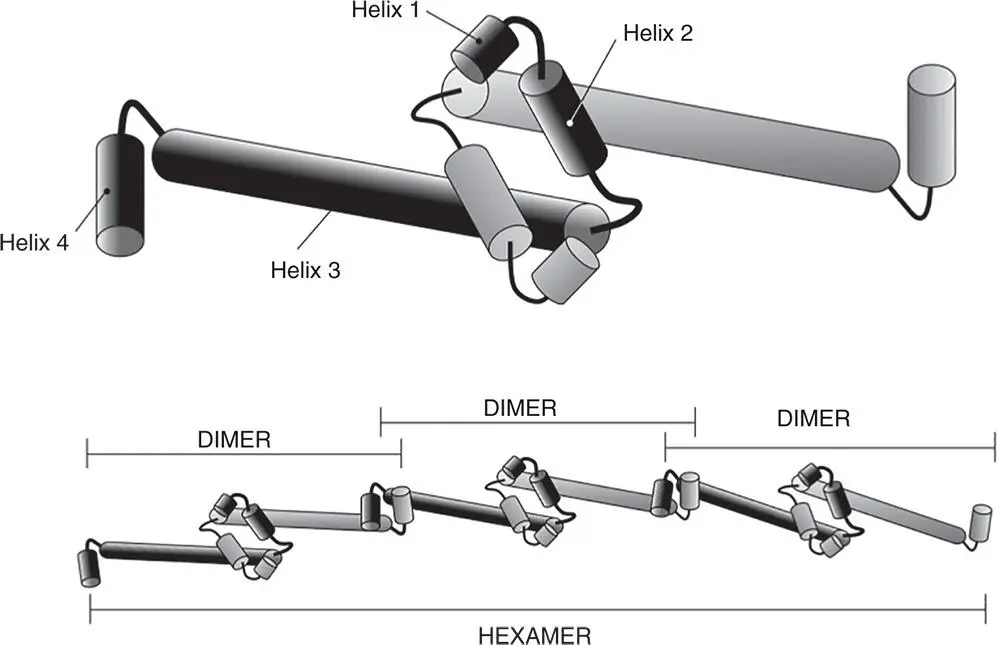
Figure 1.22The dimerisation and oligomerisation of H‐NS. The amino‐terminal domain of H‐NS is shown, with its four alpha helices represented by linked cylinders. The carboxyl terminal domain with its nucleotide binding activity and the linker region that connects this domain to the amino‐terminus are omitted. Interlinkage of alpha helices 1 and 2 of one monomer with the corresponding parts of another allows dimer formation. Linking alpha helices 4 allows two dimers to form a tetramer, and this process can continue indefinitely to produce an H‐NS polymer.
For further information, see Arold et al. (2010).
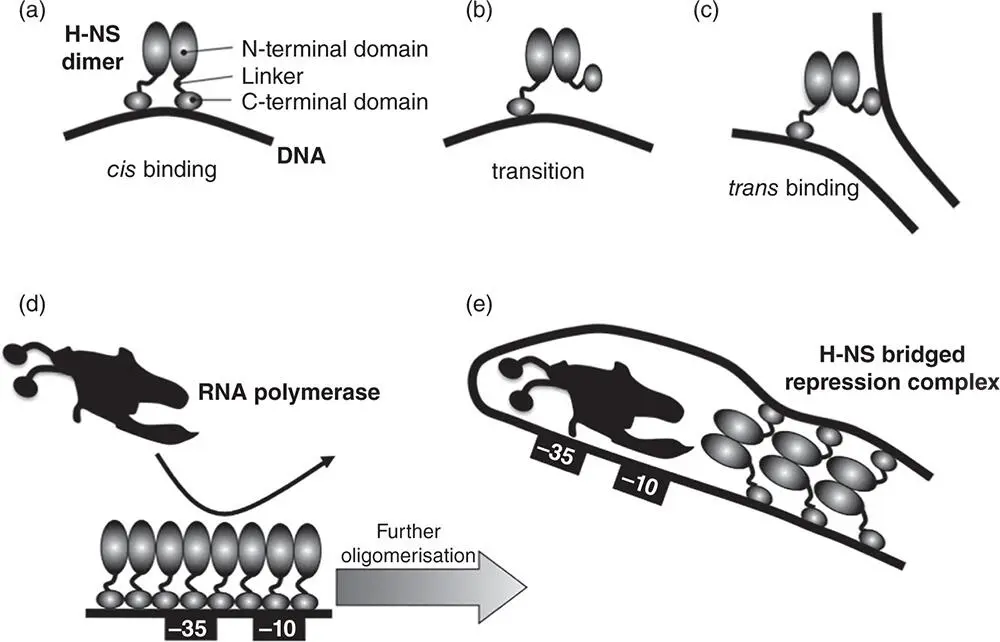
Figure 1.23The DNA‐binding modes of H‐NS. The H‐NS monomer is shown as two linked ovals with one oval representing the amino‐terminal dimerisation/oligomerisation domain and another the carboxyl‐terminal domain with the nucleic acid binding activity. The functional domains are connected by a flexible linker. (a) An individual dimer is shown in a cis ‐binding mode, with both DNA‐binding modules on the same DNA molecule. (b) It then undergoes a transition to a trans ‐binding mode in which it attaches simultaneously to two DNA molecules or to two separate sections of the same DNA molecule. (c) This trans ‐binding mode is the basis for H‐NS‐mediated DNA bridging and the transition between the two modes is toggled by Mg 2+concentrations. Either mode can silence transcription. (d) In one case polymerisation of H‐NS along DNA excludes RNA polymerase from a promoter. (e) In the other case, H‐NS bridges DNA from upstream and downstream of the transcription start site, blockading the promoter so that RNA polymerase is either excluded or becomes trapped in the initiation stages of transcription.
H‐NS displacement can be assisted by properties of the DNA to which it binds. In the genetic switch that controls the expression of the major virulence genes of Shigella flexneri , a region of curved DNA undergoes a thermally induced reorganisation that displaces H‐NS, leading to derepression of the virF regulatory gene (Prosseda et al. 2004). H‐NS may also use an internal conformational change in response to rising temperature to disrupt its transcription‐silencing complexes on DNA (Shahul Hameed et al. 2018). However, most of the anti‐H‐NS mechanisms that have been described involve roles for a second protein. Also in S. flexneri , the VirB DNA‐binding protein derepresses the promoters of the principal virulence gene operons by binding and wrapping DNA, making H‐NS binding and bridging untenable (Turner and Dorman 2007). This form of antagonism can be transferred to other H‐NS‐repressed promoters simply by appropriate placement there of the VirB‐binding site (Kane and Dorman 2011). The similarity of VirB to proteins involved in the partitioning of plasmids with protein‐dependent active partitioning systems ( Section 1.15) shows that DNA‐binding molecules can be co‐opted from other purposes to oppose the transcription silencing imposed by H‐NS (Adler et al. 1989; Kane and Dorman 2012; Turner and Dorman 2007). This suggests that simple genetic switches based on transcription silencing and anti‐silencing can arise quickly in the evolution of gene regulatory circuits.
The DNA‐binding and bridging activity of H‐NS may lend itself to nucleoid organisation as well as to transcription control (Dorman 2013; Japaridze et al. 2017). The distribution pattern of H‐NS binding sites around the chromosome appears to be periodic and genetic experiments have identified the hns gene as being important for the formation of chromosomal domain boundaries (Hardy and Cozzarelli 2005). Experiments with super resolution imaging and chromosome conformation capture have produced some data that indicate a central role for H‐NS in nucleoid architecture (Wang et al. 2011) while other 3C experiments seem to rule out a role for this protein in the organisation of long range interactions in the nucleoid (Cagliero et al. 2013). Other HiC experiments show that H‐NS promotes DNA interactions in the chromosome at short ranges, but not the long‐range interactions between H‐NS‐regulated genes and operons that were detected by Wang et al. (2011) (Lioy et al. 2018). Work with HiC in Caulobacter has implicated the high‐frequency transcription of long genes as playing a role in boundary formation and maintenance, independent of any effect of translation (Le and Laub 2016). This work suggests that the act of transcribing a long gene inhibits interactions between the DNA regions that flank that gene, causing this long gene to act as a boundary element between domains, a process that may become modulated by H-NS-mediated transcription silencing.
1.43 StpA: A Paralogue of H‐NS
The ratio of H‐NS to its genomic binding sites appears to be important for the competitive fitness of the bacterial cell. Bacteria that gain extra copies of the hns gene through the introduction of multicopy recombinant plasmids that encode this NAP gain in their capacity to replicate themselves (C.J. Dorman, unpublished). The model organisms E. coli and Salmonella typhimurium encode the StpA protein, a second H‐NS‐like molecule. It shares many features with H‐NS and has been described as an RNA chaperone (Doetsch et al. 2010). StpA can substitute for H‐NS and it can form heterodimers with its paralogue (Johansson et al. 2001; Leonard et al. 2009; Sonden and Uhlin 1996; Zhang et al. 1996 Refs). The stpA gene is expressed maximally during exponential growth, perhaps to provide an auxiliary supply of H‐NS‐like protein at a point in the growth cycle where the number of H‐NS binding sites is most numerous (Deighan et al. 2003; Free and Dorman 1997). In this context, it is interesting to note that the transcription of the hns gene is linked positively to chromosome replication (Free and Dorman 1995).
1.44 H‐NS Orthologues Encoded by Plasmids and Phage
Copies of genes encoding H‐NS‐like proteins occur naturally on plasmids, including large, self‐transmissible plasmids (Shintani et al. 2015). These molecules have A+T‐rich DNA and may impose a competitive fitness cost on bacteria that receive them in conjugation by diverting H‐NS from A+T‐rich DNA sites on the chromosome, leading to disruption of the gene expression pattern in the bacterium. By providing its own supply of H‐NS activity, the plasmid can avoid this regulatory disturbance and its associated impact on competitive fitness (Doyle et al. 2007). The R27 plasmid, originally detected in Salmonella , and more recently in S. flexneri 2a 2457T, encodes the H‐NS orthologue Sfh (Beloin et al. 2003a; Deighan et al. 2003). The two proteins exhibit considerable overlaps in their binding sites on the bacterial chromosome. When H‐NS is present, Sfh is restricted to a subset of the sites that it can occupy when H‐NS is removed by inactivation of the hns gene (Dillon et al. 2012). These observations are consistent with Sfh acting as an auxiliary to H‐NS, a role that it shares with StpA. The expression patterns of the three proteins are instructive in this regard: H‐NS is present at a constant level per chromosome throughout the growth cycle, StpA is expressed when the cells are in exponential phase, and Sfh appears at the beginning of stationary phase (Deighan et al. 2003).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Structure and Function of the Bacterial Genome»
Представляем Вашему вниманию похожие книги на «Structure and Function of the Bacterial Genome» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Structure and Function of the Bacterial Genome» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.