S. Jane Flint - Principles of Virology, Volume 2
Здесь есть возможность читать онлайн «S. Jane Flint - Principles of Virology, Volume 2» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Principles of Virology, Volume 2
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Principles of Virology, Volume 2: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Principles of Virology, Volume 2»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Volume I: Molecular Biology
Volume II: Pathogenesis and Control
Principles of Virology, Fifth Edition
Principles of Virology, Volume 2 — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Principles of Virology, Volume 2», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
In addition, populations of viruses often contain defective particles that are not capable of completing an infectious cycle. Such particles can be produced by incorporation of errors during virus genome replication or by interactions with inhibitory compounds in the environment. The ratio of infectious to defective virus particles in a preparation can be calculated by dividing the number of infectious particles (de fined using a plaque assay; Volume I, Chapter 2) by the total number of particles in a sample. Total particles, infectious and noninfectious, are classically determined using electron microscopy, although less arduous and equally quantitative strategies now exist. Some viruses, such as many bacteriophages, have a very low ratio (that is, virtually all particles are infectious), while other particle-to-PFU ratios, including those for poliovirus and some papillomaviruses, approach 1,000 or 10,000, respectively. Why these ratios differ so radically is not known, but the main point should be clear: not every virus particle that binds to a susceptible and permissive cell can induce all the steps needed to produce progeny virus particles, and even those that can may be thwarted at any step of the viral reproductive cycle.
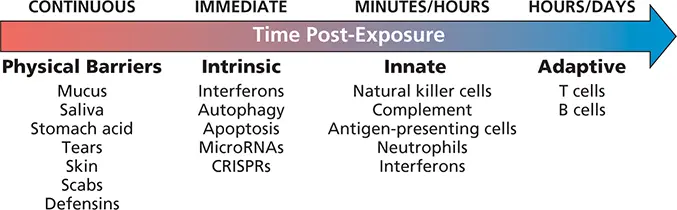
Figure 2.2 The coordinated host response to infection. In healthy individuals, anatomical and chemical barriers are in place to prevent or repel infection by microbes. When viruses successfully bypass these defenses, intrinsic responses are engaged. These responses already exist in the infected cell, poised to respond without the need for new transcription or translation. Within hours following exposure, cellular components of the innate immune response, including professional antigen-presenting cells, neutrophils, and natural killer cells, migrate to the site of infection. These infiltrating cells, as well as the infected cells themselves, produce soluble proteins (chemokines and cytokines) that serve as beacons for the eventual recruitment of T and B cells. CRISPRs, clustered regularly interspaced short palindromic repeats.
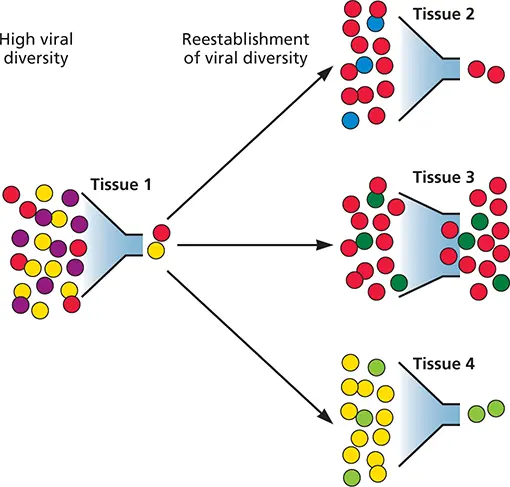
Figure 2.3 Infection seen as a series of bottlenecks. In the illustrated case, the viral population enters the host as a diverse quasispecies with sufficient titer to establish infection. After entry, the population may encounter a host barrier (bottleneck) that limits diversity. Individual members within this population (red/yellow) may overcome this bottle neck, reproducing and restoring diversity. As a result, the subsequent viral population in a given tissue may have high diversity but differ in overall consensus sequence from the initial infecting population. Note, for example, the emergence of “new” blue and green viruses not found in the original population. Certain tissues that do not impose such bottle necks may be highly permissive for viral infection (e.g., Tissue 3).
Successful Infections Must Modulate or Bypass Host Defenses
In most mammals, common sites of virus entry include the mucosal linings of the respiratory, alimentary, and urogenital tracts; the outer surfaces of the eyes (conjunctival membranes or cornea); and the skin ( Fig. 2.4). Each of these portals is equipped with anatomical or chemical features that limit viral entry and infection.
Skin
The skin is the largest organ of the body, weighing more than 9 kg (20 pounds) in an average adult. It serves obvious protective functions, but is also required for thermoregulation, control of hydration and evaporation, and integration of sensory information. The external surface of the skin, or epidermis, is composed of several layers, including a basal germinal layer of proliferating cells, a granular layer of dying cells, and an outer layer of dead, keratinized cells ( Fig. 2.5). This outermost layer is a rather literal coat of armor against viral infection: many virus particles that land on intact skin are inactivated by dehydration, acids, or other inhibitors secreted by commensal microorganisms. Some virus particles are removed from the body when dead cells slough of; many others are washed away by soap and water. However, when the integrity of the dead cell layer is compromised by cuts, abrasions, or punctures (e.g., insect bites and needle sticks), virus particles can access the rich array of live cells beneath the keratinized layer, including epithelial cells, endothelial cells, neuronal processes, and capillaries.
Examples of viruses that can gain entry via the skin include some human papillomaviruses, certain poxviruses (e.g., myxoma virus), and all tick- or mosquito-borne viruses that are transmitted by arthropod injection below the dead cell layer. Even deeper inoculation into the tissue below the dermis can occur by hypodermic needle punctures, body piercing, tattooing, or sexual contact when body fluids are mingled as a result of skin abrasions or ulcerations. Viruses that can gain entry in this manner include hepatitis B and C, human immunodeficiency virus type 1, and the herpesviruses Epstein-Barr virus and cytomegalovirus. Finally, rabies virus can be transmitted by animal bites that penetrate deep into tissue and muscle that are rich with nerve endings. Access to nerve terminals provides an opportunity for infection of motor neurons that ultimately leads to the nerve damage often associated with rabies virus infection. Superficial infections in the epidermis typically remain focalized (e.g., papillomaviruses that cause warts), whereas deeper penetration of viruses in dermal or subdermal tissues can reach nearby blood vessels, lymphatics, and neurons, conduits that enable systemic transmission ( Box 2.2).
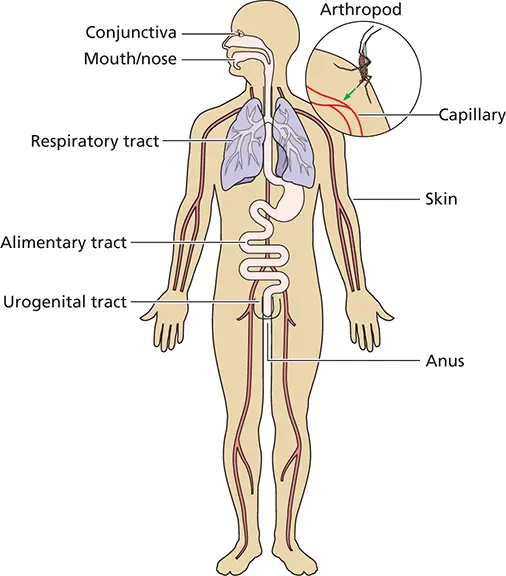
Figure 2.4 Sites of viral entry into the host. The body is covered with skin, which has a relatively impermeable (dead) outer layer of keratinocytes covering a layer of live epithelial cells rich in capillaries. Breaches in the integrity of the skin may allow viruses (or other microbes) access to this rich source of living cells. Moreover, other portals in the host, present to absorb food or release waste (mouth, urogenital tract, anus), exchange gases (respiratory tract), or interact with the environment (eyes), can also be entry points to allow access of viruses to host tissues.
The body’s response to a breach in the critical barrier formed by the skin is to make rapidly a hard, water-resistant shell over the wound, called a scab. Scabs are more than just the dermis below the site of injury drying and hardening; neutrophils and macrophages are recruited in large numbers to a wound, primarily to engulf bacteria and other pathogens that may benefit from this breach in the skin to infect the host. In addition, macrophages further aid the healing process by producing growth factors that promote cell proliferation. As the air dries the wound area, these formerly useful immune cells become part of the scab as well.
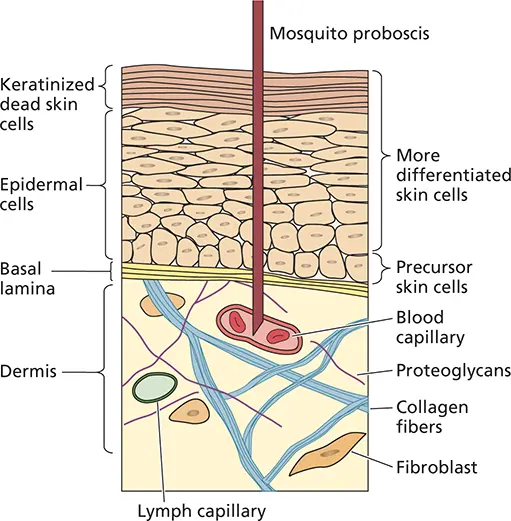
Figure 2.5 Schematic diagram of the skin. The epidermis consists of a layer of dead, keratinized cells over the live epidermal cells. Below this is the basement membrane (basal lamina). Below the basement membrane lies the dermis, which contains blood vessels, lymphatic vessels, fibroblasts, nerve endings, and macrophages. The potential depth reached by the proboscis of a mosquito taking a blood meal is shown. For more on how mosquitos spread viruses, see https://youtu.be/7wsk8a3ze80.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Principles of Virology, Volume 2»
Представляем Вашему вниманию похожие книги на «Principles of Virology, Volume 2» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.



Обсуждение, отзывы о книге «Principles of Virology, Volume 2» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.