Malcolm L. Hunter - Fundamentals of Conservation Biology
Здесь есть возможность читать онлайн «Malcolm L. Hunter - Fundamentals of Conservation Biology» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Fundamentals of Conservation Biology
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Fundamentals of Conservation Biology: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Fundamentals of Conservation Biology»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
has been a valued mainstay of the literature, serving both to introduce new students to this ever-changing topic, and to provide an essential resource for academics and researchers working in the discipline. In the decade since the publication of the third edition, concerns about humanity’s efforts to conserve the natural world have only grown deeper, as new threats to biodiversity continue to emerge.
This fourth edition has taken into account a vast new literature, and boasts nearly a thousand new references as a result. By embracing new theory and practice and documenting many examples of both conservation successes and the hard lessons of real-world “wicked” environmental problems,
remains a vital resource for biologists, conservationists, ecologists, environmentalists, and others.
Fundamentals of Conservation Biology — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Fundamentals of Conservation Biology», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Last, geneticists often use the heterozygosity index to estimate how much of a species’ total genetic diversity (H t) is due to genetic diversity within the populations that compose the species (H s) versus how much is due to variability among those populations (D st) (Nei and Kumar 2000). Mathematically, this can be expressed as H t= H s+ D st. (This concept is often expressed with different but related formulas, but the basic idea is the same: partitioning the overall variability that exists in a species within and between the populations that comprise it.) This may seem an arcane concept but it is quite useful. If a species has a relatively high D st, then it is necessary to maintain many different populations to maintain the species’ overall genetic diversity. For example, many salamanders have extraordinarily high values of D st, which emphasizes the importance of preserving many populations that comprise a species to “capture” the genetic diversity that makes up the species (Tilley 2016). Alternatively, if most of the species’ genetic diversity exists within every population (i.e. H sis relatively high), then it is less critical to maintain many different populations ( Fig. 5.4). This is the case, for example, for many (but not all) birds, which fly and disperse well, intermixing their populations and thereby lowering D st(e.g. for endangered piping plovers, Haig and Oring 1988). These considerations are often important for people who manage populations of endangered species, and we will return to in Chapter 13, “Managing Populations.”
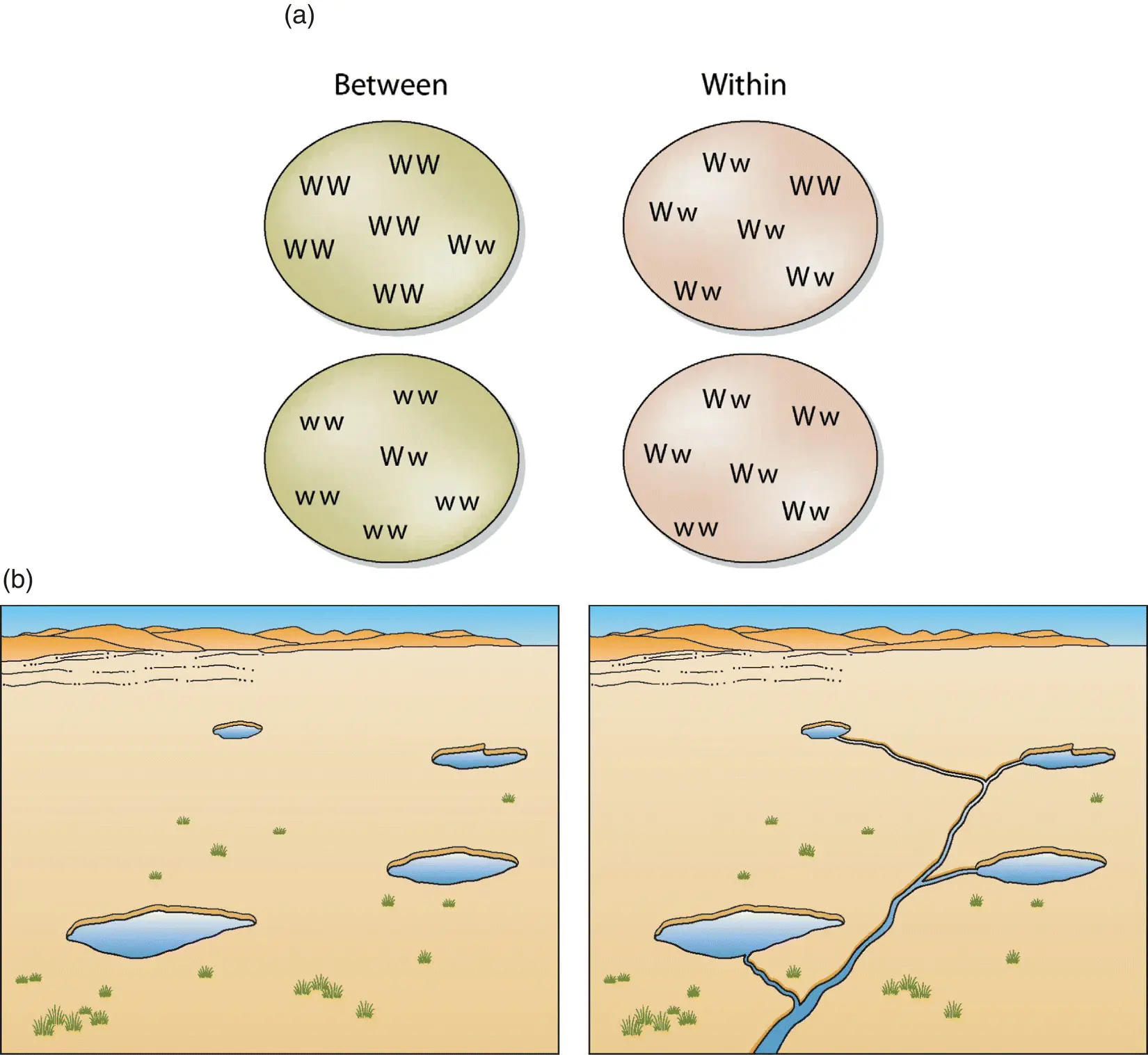
Figure 5.4 Genetic diversity is partitioned within versus among populations to varying degrees with important implications for conservation strategies. First we tackle this conceptually (a). In the first case (“between”), the two alleles present (“W” or “w”) are each sequestered into different populations. Here conserving genetic diversity can be accomplished only by protecting both populations. In the second case (“within”), each population has both alleles present, and protecting a single population captures all the diversity present. In more practical terms (b), desert fishes living in isolated springs ( left ) will likely have higher genetic variability among populations (higher D st) than desert fishes in which populations are connected by streams through which the fishes can disperse ( right ).
Quantitative Variation
So‐called “qualitative variation,” such as allele frequencies in populations, tells us much about how species are organized and their history, but the key traits that most determine fitness are in fact “quantitative” characters, such as height, weight, litter size, seed set, survival rate, etc. (Frankham et al. 2009). Such traits vary continuously because they are polygenic (controlled by many genes) and affected by the environment as well. Adaptive evolution results from changes in quantitative traits, so scientists study quantitative traits because they tell us much about the capacity of a population to evolve in response to environmental change (Storfer 1996). As an example, consider coral reefs, which are threatened by increasing ocean temperatures. Acute temperature increases are stressful for corals, but gradual temperature changes can result in adaptation depending on how heat tolerant a coral is. Heat tolerance is a “quantitative” trait because it varies gradually, relatively higher or lower from individual to individual (versus if it were “qualitative” = heat tolerant or heat intolerant) and is an inherited trait. This natural and quantitative variation in temperature tolerance may facilitate rapid adaptation among corals as our oceans warm (Dixon et al. 2015) with some researchers even suggesting transplanting more heat tolerant coral individuals to reefs that are warming rapidly. As another example, the endangered, annual plant known as the spinster’s blue‐eyed Mary, grows on and off serpentine soils, which have high concentrations of some toxic minerals and low concentrations of some essential nutrients. Plants from serpentine habitats grow best in serpentine habitats, and plants from nonserpentine habitats do best in nonserpentine habitats ( Fig. 5.5) even though they are from populations as little as 100 m apart. Restoration efforts need to focus on collecting locally adapted seeds, as using nonserpentine seed sources in serpentine areas may lead to failure (Wright et al. 2006).
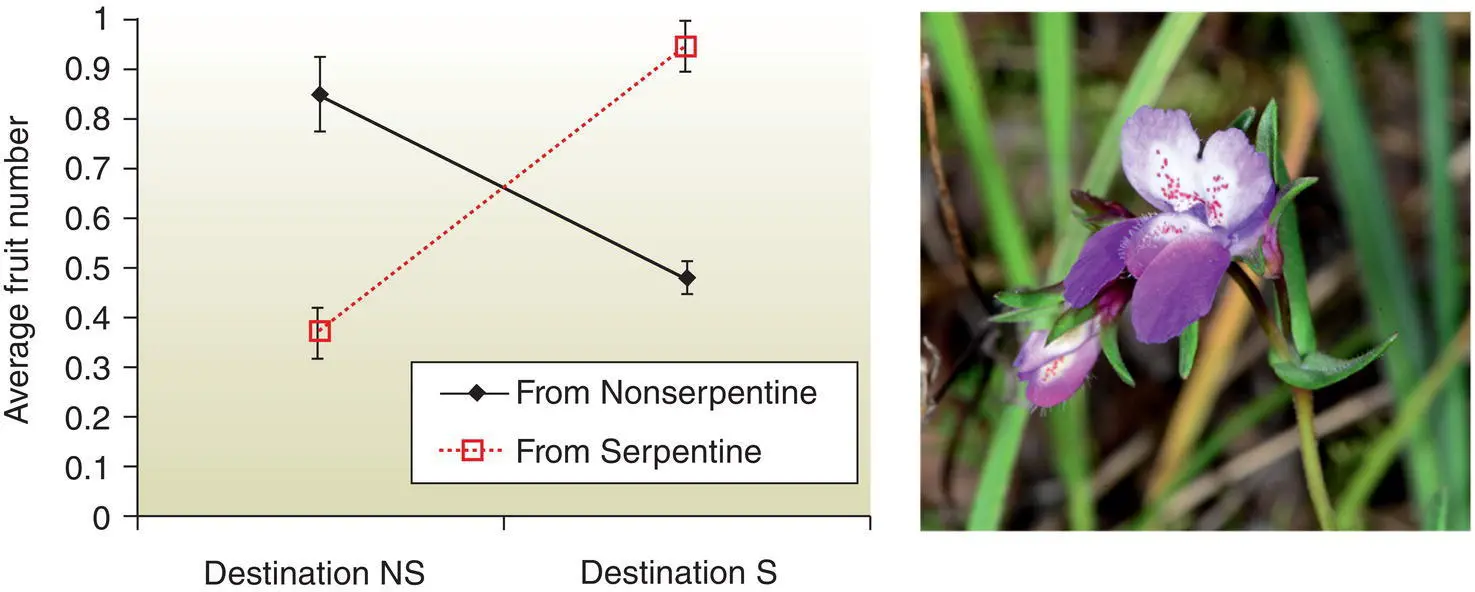
Figure 5.5 The native annual plant, Collinsia sparsiflora , grows on [“S”] and off [“NS”] serpentine soils with plants from serpentine habitats growing best in serpentine habitats, and plants from nonserpentine habitats doing best in nonserpentine habitats.
(Adapted from Wright et al. 2006 [left]; Don Loarie [right])
The Importance of Genetic Diversity
We have touched on the importance of genetic diversity already. To really understand the importance of genetic diversity, it is useful to think of genes as units of information rather than tangible things. As tiny aggregations of carbon, hydrogen, oxygen, nitrogen, and some other common elements, genes have little value in and of themselves. Indeed, when DNA is extracted from any organism and concentrated in the bottom of a test tube it appears as a small, sticky, gray, and rather unattractive blob. As sources of information, however, genes are clearly essential; they shape the synthesis of the biochemicals that control cellular activity and, ultimately, all biological activity and form. The capacity for genes to encode this information is stupendous; a typical mammal might have 100,000 genetic loci.
Of course, most of this wealth of genetic diversity is encapsulated in the diversity of species and their interspecific genetic differences. The key issue to address here is the distribution and diversity of alleles that characterize a species. Why is it important to maintain different versions of the same gene and, in many circumstances, to have them well distributed in a population dominated by heterozygotes rather than homozygotes? There are three basic answers: evolutionary potential, loss of fitness, and utilitarian values.
Evolutionary Potential
A key requisite for natural selection is genetic‐based variability in the fitness of individuals; that is, some individuals must be more likely to survive and reproduce than others. If every individual were genetically identical and only chance determined which ones left progeny, then populations would change erratically through time, if at all. If they are to persist, however, populations must change as their environment changes, which environments everywhere are now doing rapidly (see Chapter 6). Of course, the physical world has always changed as continents drift around the globe, mountains rise and erode, oceanic currents and jet streams shift paths, and the planet’s orbit around the sun varies. The biological world also changes as species evolve, become extinct, and shift their geographic ranges, coming into contact with new species that may be predators, prey pathogens, or competitors. Changes have been particularly dramatic during the last few decades as human populations and their technological capabilities have grown and profoundly altered the conditions for evolution in most species. To put it more directly: humans are now the central organizing reality around which all nonhuman life will evolve. To some degree, all species must respond to the environmental changes we are wreaking almost everywhere if they are to survive. And they need genetic diversity to do so.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Fundamentals of Conservation Biology»
Представляем Вашему вниманию похожие книги на «Fundamentals of Conservation Biology» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Fundamentals of Conservation Biology» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.